






异特龙属
词源
在1877年,奥塞内尔·查利斯·马什最初为异特龙命名时,他认为这种恐龙的脊椎构造独特,和当时已知的其他恐龙有异,所以取名为异特龙(Allosaurus),allos/αλλος在古希腊文里意为“奇特的”或“不同的”,而saurus/σαυρος意为“蜥蜴”。因此异特龙意为“奇特的蜥蜴”。
但是,在翻译异特龙的中文译名时出了一些误会,有人认为该词源是拉丁文中“跳跃”的意思,所以误把它译为跃龙。但异特龙的学名应意为“奇特的蜥蜴”,与“跳跃的蜥蜴”没有关系,所以还是应该称之为异特龙。
形态

异特龙与人类的体型相比。由大到小分别是:依潘龙、AMNH680、脆弱异特龙、“大艾尔”。
异特龙是种典型的大型兽脚类恐龙,拥有大型头颅骨、粗壮的颈部、长尾巴、以及缩短的前肢。脆弱异特龙是最著名的种,平均身长为8.5米,而最大型的异特龙标本(编号AMNH 680)的身长估计为9.7米,体重为2.3公吨。在1976年,詹姆斯·麦迪逊(James Madsen)的异特龙专题论文中,他提出异特龙的身长最大值为12到13米。如同其他的恐龙,异特龙的体重估计值也有争议,自从1980年代以来,成年异特龙的体重估计值,已有1,500公斤、1,000到4,000公斤、以及1,010公斤等不同的数据。在近年,莫里逊组专家约翰·福斯特(John Foster)提出,大型的成年脆弱异特龙的体重为1,000公斤,但根据他所测量、参考的股骨,合理的估计值应约700公斤。在2009年,其他古动物学家以亚成年的“大艾尔”作为研究对象,利用电脑模型计算出“大艾尔”的体重接近1,500公斤,若计入参数变化,大概是1,400到2,000公斤之间。
目前有数个巨型标本被归类于异特龙属,但可能事实上属于其他恐龙。异特龙的近亲食蜥王龙(编号OMNH 1708)身长可能有10.9米,曾被归类于异特龙的一种,巨异特龙(A. maximus),最近的研究多认为它们是个别的属。另一个可能属于异特龙的标本(编号AMNH 5767),曾长期被归类于依潘龙(Epanterias),身长为12.1米。近年在新墨西哥州莫里逊组的彼得森采石场,发现一个大型的异特龙科部分骨骼,可能是食蜥王龙的第二个标本。

异特龙的颅骨,位于华盛顿国立自然历史博物馆

异特龙的头部重建模型
头颅骨
在兽脚亚目之中,异特龙的头颅骨、牙齿与身体的比例适中。葛瑞格利·保罗(Gregory S. Paul)依据一个长度为84.5公分的异特龙头颅骨,估计该个体的身长为7.9米。每块前上颌骨各有5颗牙齿,牙齿的横剖面呈D形,而每块上颌骨约有14到17颗牙齿;异特龙的牙齿数量与骨头大小并不呈正比。而每块齿骨约有14到17颗牙齿,平均数量为16颗。越往嘴部深处,牙齿就越短、狭窄、弯曲。异特龙的牙齿都为锯齿状。这些牙齿很容易脱落,所以它们会不断地生长、替代,并成为常发现的化石 。
异特龙的眼睛上方拥有一对角冠,由延伸的泪骨所构成。角冠的形状与大小随者个体而不同。鼻骨的上方也有一对低矮的棱脊,并沿者鼻骨,连接到眼睛上的角冠。这些角冠可能覆盖者角质,并具有不同的功能,例如:替眼睛遮辟阳光、视觉展示物、以及物种内的打斗行为(问题是它们非常脆弱)。头颅骨后上方也有一个棱脊,可供肌肉附着,这特征也可见于暴龙科动物。
泪骨内侧有凹处,可能内藏腺体,例如盐腺。上颌骨内侧有凹陷处,发展的比基础兽脚类恐龙的鼻窦还好,例如角鼻龙与马什龙;这些凹陷处可能与嗅觉器官有关,例如犁鼻器。异特龙脑壳顶部较薄,可能为促进脑部的体温调节。
异特龙的头颅骨与下颌是由个别的骨头所组成,而骨头之间有可活动关节。例如下颌的前半部与后半部可往外弯曲,增加骨头间的空隙,因此可以吞下较大的食物。脑壳与额骨之间可能也有类似的关节。
骨骼

异特龙的骨骼重建。位于圣地牙哥自然历史博物馆
异特龙拥有9节颈椎、14节背椎、5节支撑臀部的荐椎。尾椎的数量不明,可能随者个体大小而不同;詹姆斯·麦迪逊估计异特龙有接近50节尾椎,而葛瑞格利·保罗认为这个数量过多,提出应该不超过45节。颈椎与前段背椎有中空区域,这种空间也可见于现代鸟类,被认为具有类似鸟类的气囊系统,使用于呼吸作用上。异特龙的肋骨宽广,形成桶状胸腔,与较原始的兽脚类(例如角鼻龙)不同。异特龙也具有腹肋(一种真皮骨),但不常被发现,可能只有稍微的骨化。在一个已公布标本中,这些腹肋被发现生前曾受过伤。有一个叉骨被保存下来,但直到1996年才被确认出来;在一些案例中,叉骨与腹肋则被混淆。主要的臀部骨头肠骨巨大,耻骨有个明显的尾端,可能作为肌肉附着处,以及身体躺在地面时的支撑物。在1976年,麦迪逊发现克利夫兰劳埃德恐龙采石场所发现的异特龙化石,有接近一半个体的两个耻骨上端,并未互相固定者。由于这个特征与体型无关,因此麦迪逊认为这是种两性异形,雌性个体的耻骨上端没有互相固定者,可使产卵时更为顺利。然而,这个理论并未引起进一步的讨论。

脆弱异特龙的手掌与指爪
与后肢相比,异特龙的前肢相当短,约是后肢长度的35%。每个手部具有三根手指,以及大型、大幅弯曲的指爪。异特龙的前肢强壮,与其他的兽脚类恐龙相比,它们的前肢适合抓握一定距离内的猎物,或是将猎物拉近。前臂稍短于上臂,肱骨与尺骨的比例为1:1.2。手腕具有类似半新月形的腕骨,手盗龙类的腕骨更为接近半新月形。异特龙的三根手指中,内侧第一根手指是最长的。指爪的状态显示手指可能用来钩住东西。
与暴龙科相比,异特龙的腿部较短,不像暴龙科适合奔跑,而且趾爪较原始,类似早期兽脚类恐龙的蹄状趾爪。每个脚部具有三个巨大趾爪,可以承受它们的巨大重量;第四趾萎缩成为上爪,麦迪逊认为幼年个体的上爪具有挖握功能。异特龙被认为具有残余的第五跖骨,可能作为跟腱与脚部的夹层使用。
分类

破碎异特龙的想像图
异特龙属于异特龙科,异特龙科是一个大型兽脚亚目的演化支。奥塞内尔·查利斯·马什在1878年建立异特龙科;但在1970年代以前,异特龙科很少被使用,取而代之的是斑龙科,但斑龙科长期以来被当作大型兽脚类恐龙的“未归类物种集中地”。在1976年麦迪逊公布他的专题论文以前,跟异特龙相同时期被命名的腔躯龙,常被当作异特龙的同义词,而且更常被使用。数个研究人员曾在他们的著作中使用斑龙科,而不采用异特龙科,包含:吉尔摩尔、休尼博士、罗默博士、罗德尼·史提尔、以及亚历克·沃克。
在麦迪逊公布他的具影响力专题论文之后,异特龙科开始普遍使用,但没有被明确的定义。一些半科学性质的著作将异特龙科使用于不同的大型兽脚类恐龙,通常包含那些比斑龙科更大型、更著名的物种。在兽脚亚目中,常被认为是异特龙近亲的物种,包含:印度龙、皮亚尼兹基龙、皮尔逖龙、永川龙、高棘龙、吉兰泰龙、巧鳄龙、史托龙、以及四川龙。随者相关恐龙的研究增加,兽脚类恐龙开始变得多样性。而亲缘分支分类法研究则显示,上述原本被认为是异特龙近亲的物种,都不属于异特龙科,只有少数属被归类于相近的科,例如:高棘龙、永川龙。
异特龙科是肉食龙下目的四个科之一,其他三个分别为新猎龙科、鲨齿龙科与中华盗龙科。在1988年,葛瑞格利·保罗提出异特龙科演化为暴龙科,因此成为并系群;但这个看法已遭到否定,暴龙科已经被归类为另一群兽脚类演化支,虚骨龙类。异特龙科是肉食龙下目中最少属的一科,在大多数的近期研究中,除了异特龙属以外,目前只有食蜥王龙以及一个发现于法国的未命名异特龙超科恐龙被认为是可能的有效属。依潘龙(Epanterias)是另一个可能的有效属,但依潘龙与食蜥王龙有可能是异特龙属的大型个体。最近的研究则保留了食蜥王龙,而将依潘龙归类于异特龙的一个种。
发现和命名
早期的发现与研究
早在1869年,科罗拉多州格兰比郡附近的中央公园当地居民将一个恐龙化石误认为是一个“马蹄化石”,并交给了费迪南德·范迪威尔·海登(Ferdinand Vandiveer Hayden);该地可能属于莫里逊组。海登将这标本交给约瑟夫·莱迪,莱迪发现这个“马蹄化石”其实是半节尾椎骨。约瑟夫·莱迪起初暂时将它归类为欧洲杂肋龙的一个种(Poicilopleurodon valens)。后来在1873年,莱迪将它建立为一个新的属,腔躯龙(Antrodemus)。

AMNH 5753重建图,正在吞食迷惑龙的尸体。由查尔斯·耐特所绘制。

现今的AMNH 5753重建模型,依照查尔斯·耐特的图画重建。
异特龙的正模标本(编号YPM 1930)被发现于科罗拉多州卡农城北方的花园公园,由一小群破碎骨头所构成,包含三节脊椎、一个肋骨碎片、一颗牙齿、一个趾骨、以及右肱骨主干部分;而右肱骨最常被后来的研究提及。在1877年,奥塞内尔·查利斯·马什根据这些化石,把这种生物定名为异特龙属,并把其模式种正式命名为脆弱异特龙(Allosaurus fragilis)。种名fragilis来自拉丁语,意即“脆弱的”,是指它们脊椎骨的轻盈特点。
在18世纪后期的化石战争期间,马什与爱德华·德林克·科普之间有过激烈的竞争。两人曾根据许多零散、相似的化石建立了数个属,但之后被证明属于异特龙,使得异特龙的发现与早期研究非常复杂。两人所建立的名称包含:马什命名的Creosaurus(意为“肌肉蜥蜴”)、Labrosaurus(意为“贪吃的蜥蜴”)以及科普命名的依潘龙(Epanterias,意为“拱起的”)。
在竞争的过程中,科普与马什并没有持续地进行他们与他们下属的挖掘工作。举例而言,本杰明·福兰克林·马奇(Benjamin Franklin Mudge)在科罗拉多州花园公园发现异特龙的正模标本后,马什便转向怀俄明州进行新的挖掘工作;而在1883年,M. P. Felch重新开始花园公园的挖掘工作后,却发现了一个几乎完整的异特龙化石,以及数个部分骨骸。另外,科普的一个挖掘工人H. F. Hubbell,在1879年于怀俄明州的科摩崖发现了一个异特龙标本(编号AMNH 5753),但他没有提到化石的完整程度,所以科普从未看过这个化石。在1903年,科普死后数年,这个标本被发现是当时最完整的兽脚类恐龙之一,并在1908年开始展览。在一个由查尔斯·耐特(Charles R. Knight)所绘制的图画中,编号AMNH 5753的异特龙跨越在一只迷惑龙身上,吞食着迷惑龙的尸体。虽然这是第一次将兽脚类恐龙描绘成站立姿态,但当时并没有科学证据可以支持。
异特龙名称的重复使得研究变得复杂,并随着马什与科普的竞争而恶化。在当时便有一些科学家,例如塞缪尔·温德尔·威利斯顿,提出有太多相关名称被重复建立。在1901年,威利斯顿便指出马什自己也无法分辨出异特龙与Creosaurus的差异。查尔斯·怀特尼·吉尔摩尔在1920年尝试将这些复杂的名称整理、归类。吉尔摩尔认为莱迪用来命名腔躯龙的尾椎,其实跟异特龙的尾椎一样;因为腔躯龙较早命名,所以取代异特龙。在接下来的50年,腔躯龙取代异特龙,成为正式的名称,直到詹姆斯·麦迪逊在克利夫兰劳埃德采石场发现新的异特龙化石,并出腔躯龙的化石材料无法鉴定,发现地点也不清楚,应该采用异特龙这名称。腔躯龙目前为非正式的用法,只用在区分吉尔摩尔与麦迪逊两人重建的不同形态头颅骨。

一个异特龙的早期骨架模型,位于新西兰基督城坎特伯里博物馆。现在认为异特龙是采用水平的步态
克利夫兰劳埃德
早在1927年开始,犹他州艾麦里县的克利夫兰劳埃德恐龙采石场便有了零散的发现,威廉·斯托克斯(William J. Stokes)在1945年于《科学》杂志描述了这个采石场,但直到1960年代,才开始了大型的挖掘计划。在1960年到1965年期间,在接近40个机构的合力挖掘之下,该采石场出土了数千块骨头。克利夫兰劳埃德采石场的著名原因包含:大部分骨头都属于脆弱异特龙,例如在2006年,73个出土的恐龙个体中,至少有46个属于脆弱异特龙;这些化石的关节脱落,而且互相混合;将近有10多个科学研究讨论了该地点的化石埋葬状况,形成不同、互相矛盾的解释。对于该地的成因,包含大群动物深陷在泥泞之中、干旱导致大群动物困在水洼之中。无论正确的成因为何,当地发现的大量异特龙化石使得科学家可以详细地研究它们,使得异特龙成为研究最多的兽脚类恐龙之一。该地所出土的异特龙化石几乎包含各种年龄与大小,身长范围从1米到12米。
80年代以来的研究
自从麦迪逊的专题论文公布以来,已有许多研究讨论异特龙的古生物学以及古生态学。这些研究涵盖了异特龙的骨骼差异、生长模式、头颅骨结构、猎食模式、脑部构造、以及群居生活与亲代养育的可能性。早期异特龙标本的重新研究、在葡萄牙新发现的化石、以及数个非常完整的标本也有助于对于异特龙的了解。在2000年代早期,波兰古生物学家Gerard Gierliński在波兰Bałtów发现一个足迹化石,被归类于异特龙。

“大艾尔二号”骨架模型,位于休士顿自然科学博物馆
大艾尔
在1991年发现的“大艾尔”标本(编号MOR 593),是最著名的异特龙化石之一。大艾尔是个相当完整的天然状态标本,接近95%完整度,身长约8米。“大艾尔”最初是由卡比·希伯(Kirby Siber)所率领的瑞士团队发现于怀俄明州的比格霍恩县,并由怀俄明大学地理博物馆与洛矶山博物馆共同挖掘出土。因为该化石相当完整,因此取了个昵称“大艾尔”。发现大艾尔的瑞士团队后来在同一地点发现另一异特龙化石,并取名为“大艾尔二号”,是到目前为止,保存状态最好的异特龙化石。
“大艾尔”的体型小于脆弱异特龙的平均大小,可能是个亚成年体,经估计只成长了87%,或是另一较小的种。“大艾尔”由布伦特·布莱特豪普特(Brent Breithaupt)是在1996年所叙述。其中有19个骨头是断裂的,呈现出感染的迹象,这可能导致“大艾尔”的死亡。这些感染的骨头包含五个肋骨、五节脊椎、四个脚部骨头;数个骨头则具有骨髓炎。右脚上的感染与伤口可能影响了大艾尔的行走,从步态的改变研判,右脚的伤口可能感染了另一只脚。
BBC《与恐龙共舞》(Walking with Dinosaurs)的特别节目《异特龙之谜》(Ballad of Big Al)便是以“大艾尔”作为主角。
种

异特龙的头颅骨(编号DINO 11541),发现于恐龙国家纪念公园,仍位于母岩
现在仍不确定异特龙属究竟有多少个种类。自从1988年以来,已有七个种被视为是可能的有效种,例如:模式种脆弱异特龙(A. fragilis)、合异特龙(A. amplexus)、残暴异特龙(A. atrox)、欧洲异特龙(A. europaeus)、巨异特龙(A. maximus)、坦达格鲁异特龙(A. tendagurensis)、以及尚未有正式叙述的詹氏异特龙("A. jimmadseni"),但其中只有少数种普遍被视为有效的种。此外,还有至少10个疑名或未叙述的无资格名称曾被归类于异特龙属。在最近期的基础坚尾龙类兽脚类恐龙的研究中,只有脆弱异特龙、詹氏异特龙(被列为未命名种)、以及坦达格鲁异特龙被承认是可能的有效种;而合异特龙(即为依潘龙)、残暴异特龙被视为是脆弱异特龙的异名,欧洲异特龙并没被承认,巨异特龙则被建立为食蜥王龙。
合异特龙、残暴异特龙、脆弱异特龙、詹氏异特龙、巨异特龙的化石都来自于莫里逊组,莫里逊组散布于美国的科罗拉多州、蒙大拿州、新墨西哥州、奥克拉荷马州、南达科他州、犹他州、以及怀俄明州,年代为上侏罗纪的启莫里阶到提通阶。脆弱异特龙是该地层最常发现、最著名的异特龙化石,目前已发现至少60个个体的化石。自从80年代以来,有研究怀疑莫里逊组是否存在者另一种异特龙,残暴异特龙;近年的研究仍维持莫里逊组只有脆弱异特龙的看法,而该地层所发现的不同异特龙化石间的差异,被认为是不同个体间的变化。欧洲异特龙的化石发现于葡萄牙劳尔哈组的波多诺伏,年代为启莫里阶,可能与脆弱异特龙是同一种动物。坦达格鲁异特龙被发现于坦桑尼亚姆特瓦拉区的坦达格鲁,年代也为启莫里阶。目前大部分的研究把坦达格鲁异特龙视为异特龙的有效种,但它有可能是一种基础坚尾龙类恐龙,或仅是兽脚亚目的一个疑名,虽然归类仍不确定,坦达格鲁异特龙是种大型兽脚类恐龙,身长可能约10米,体重约有2.5公吨。

Creosaurus atrox的正模标本素描图
异特龙的可能异名有腔躯龙、Creosaurus、Labrosaurus、以及依潘龙(Epanterias)。而异特龙大部分的种是根据零碎的化石来命名,许多被认为是破碎异特龙的异名,或者是被错误归类到异特龙属。其中一个例子是Labrosaurus ferox,是由马什在1884年所命名,但他是根据一个外形奇怪的部分下颌,而颌部前端的齿列有个明显的缺口,颌部后段则延展并外翻。后来的研究人员提出这个这个下颌骨头具有病状,应该是该动物生前受伤的痕迹,而后段变形的部分原因为石膏重建的后果。这个化石现在被认为属于脆弱异特龙。被认为属于异特龙的化石散布于全世界,包含澳洲、西伯利亚、以及瑞士,但这些化石被认为其他的恐龙。
脆弱异特龙(A. fragilis)的模式标本(编号YPM 1930)相当破碎,包含数节不完整脊椎、四肢骨头的碎片、肋骨碎片、以及一颗牙齿。数名古生物学家认为这个化石的状态破碎,不足以成为物种建立时、化石归类时的比对对象,因此提出脆弱异特龙是个疑名,甚至异特龙属本身也是个疑名。在2010年,葛瑞格利·保罗(Gregory S. Paul)、肯尼思·卡彭特(Kenneth Carpenter)向国际动物学术语命名委员会提出请求,要求将脆弱异特龙的模式标本改为编号USNM4734标本。这个要求目前还在审议中。
古生态学
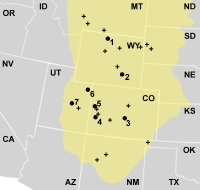
莫里逊组(黄色区块)中的著名异特龙化石挖掘地点。1.“大艾尔”2.科摩崖3.花园公园4.干梅萨采石场5.大章克申6.恐龙国家保护区7.克利夫兰劳埃德。标“+”者为其他的挖掘地点。
异特龙是美国西部莫里逊组地层中最常见的大型兽脚亚目化石,它们位在该地食物链的最上层。莫里逊组被认为是半干旱的环境,具有明显的雨季与干季,地形为平坦的泛滥平原。该地层的植被是由针叶树、树蕨、蕨类所构成的水边树林,以及由蕨类、少数短叶杉(类似南洋杉属)所构成的疏林莽原。
莫里逊组是个富含化石的挖掘地点,该地曾发现的植物化石包含:绿藻、真菌、苔藓、木贼、蕨类、苏铁、银杏、以及数科针叶树;其他的动物化石则包含:双壳纲、蜗牛、辐鳍鱼纲、青蛙、蝾螈、乌龟、喙头蜥目、蜥蜴、陆生与水生的鳄形超目、数种翼龙目、大量的恐龙、以及早期的哺乳类,例如:柱齿兽目、多瘤齿目、对齿兽目、三尖齿兽目。莫里逊组所发现的恐龙化石则包含了兽脚亚目的角鼻龙、嗜鸟龙、蛮龙,蜥脚下目的迷惑龙、腕龙、圆顶龙、梁龙,鸟臀目的弯龙、橡树龙、剑龙。在异特龙的化石发现地点,经常发现迷惑龙、圆顶龙、梁龙、剑龙的化石。
在犹他州的克利夫兰劳埃德采石场,有一个数量众多的化石层。这个化石层包含超过12,000个骨头,来自于73个个体。大部分属于异特龙化石,但也有其他恐龙化石,例如剑龙与角鼻龙。如此众多的动物化石为何集中于同一地点,目前仍不清楚。而且肉食性恐龙的比例大于草食性恐龙,这种状况非常的少见。这个化石层被解释成群体猎食所造成的,但很难证实。另一个可能性是克利夫兰劳埃德恐龙采石场在过去是个“掠食动物陷阱”,类似拉布雷亚沥青坑,造成大量的掠食动物陷入无法挣脱的沉积层中。
葡萄牙所发现异特龙的地层,年代属于晚侏罗纪,被认为环境类似莫里逊组,但受到海洋的影响较多。这个葡萄牙地层也发现了许多莫里逊组的恐龙,主要有异特龙、角鼻龙、蛮龙、迷惑龙,或者是相近物种,例如腕龙的近亲葡萄牙巨龙、弯龙的近亲龙爪龙。

莫里逊组的恐龙国家保护区
异特龙与其他侏罗纪兽脚类恐龙共同生存在莫里逊组与葡萄牙,包含:角鼻龙、体型较大的蛮龙,这点很不寻常。根据生理结构与化石位置,它们三者似乎拥有不同的生态位。角鼻龙与蛮龙可能活跃于河道附近,它们的身体较瘦、较低矮,可使它们躲藏在森林或矮树丛中;异特龙的腿部较长、速度较快,但它们在森林或矮树丛中较不灵活,因此可能活跃于干燥的泛滥平原中。目前对于角鼻龙的研究比蛮龙还深入。角鼻龙与异特龙的差别在于,它们的头颅骨较高、较狭窄,而牙齿较大、较宽广。在一个异特龙的耻骨末端发现了另一个兽脚类恐龙的齿痕,可能是由角鼻龙或蛮龙所留下的,这显示异特龙可能也是其他兽脚类恐龙的食物来源选择之一。耻骨是位于腹部下缘,夹处两腿之间,处于身体最庞大的地方之一。显示这只异特龙是在死后,尸体被其他恐龙所吞食。
古生物学
生长模式
异特龙的大量化石几乎涵盖了所有的年龄层,这使得科学家们可以研究异特龙的生长模式与年龄上限。在科罗拉多州发现的一堆压碎的蛋化石,可能属于异特龙,这是目前所发现最年幼的异特龙化石。根据四肢骨头的组织学分析,异特龙的年龄上限大约为22到28岁,相当于其他大型兽脚类恐龙(例如暴龙)。异特龙的最高生长率大约发生在15岁左右,一年可以增加150公斤的体重。

幼年詹氏异特龙的骨架模型 - 北美洲古生物博物馆
目前已在一个出土于克利夫兰劳埃德的异特龙胫骨,发现了骨髓骨组织。除了异特龙以外,腱龙与暴龙也发现了骨髓骨。骨髓骨目前只存在于产卵的雌性鸟类身上,骨髓骨富含钙,可用来制造蛋壳的。异特龙的骨髓骨组织,显示该个体是雌性的,而且正在繁衍期中。这个雌性异特龙估计是在10岁时死亡,从此显示异特龙在完全成长前,就已达到性成熟。
一个具有几乎完整后肢的幼年异特龙标本,显示幼年个体的后肢比例较成年个体长,而且后肢下半部(小腿与脚部)长于大腿部分。这些差别显示年轻异特龙的移动速度较快,并具有不同于成年个体的猎食方式,例如追赶小型猎物,而成年个体则改采伏击方式捕食大型的猎物。随者异特龙的成长,它们的股骨变得更厚、更宽,而横剖面变得较不圆形,随者肌肉附着点的改变,肌肉相对更短,腿部的成长减缓。这些改变显示幼年异特龙的腿部,承受的应力较成年异特龙小,幼年异特龙可能以更规律的速度前进。相对地,异特龙的头颅骨在成长阶段不会改变比例,头部各部位的相对比例不会大幅改变。
进食方式

异特龙的攻击方式。根据巴克与雷菲尔德等人的理论而重建。
异特龙被认为是种主动攻击的大型掠食动物。根据蜥脚类恐龙骨头上的异特龙齿痕,以及与蜥脚类化石一起发现的零散异特龙牙齿来判断,异特龙可能以蜥脚类恐龙为猎食对象,或是搜寻它们的尸体为食。另外有明确证据显示异特龙曾经攻击过剑龙,例如一个异特龙的尾椎上有个部分痊愈的伤口,这个被刺穿伤口的形状符合剑龙的尾刺;另外,在一个剑龙的颈部骨板上有个U形的伤口,与异特龙的嘴部形状符合。在1988年,葛瑞格利·保罗(Gregory Paul)提出异特龙不可能以蜥脚类恐龙为食,除非采取群体方式猎食;因为异特龙的头部大小属中型、牙齿相对较小,体型也无法与同时代的大型蜥脚类恐龙相比。另一个可能则是异特龙以幼年蜥脚类恐龙为猎食对象,而不猎食完全成长的蜥脚类恐龙。1990年代与2000年代的研究可能解答了这个问题。罗伯特·巴克(Robert T. Bakker)将异特龙与一些生存于新生代肉食性哺乳类相比,发现类似的适应演化,例如:颌部肌肉的缩小、颈部肌肉的增大、以及将颌部左右撑开的能力。虽然异特龙的牙齿并非如这些哺乳类呈刀刃状,巴克提出了另一种异特龙的攻击方式:上颌的短牙齿会形成类似锯子的小型锯齿表面,可切入猎物肉体。这种型态的颌部可使异特龙采取撕咬方式攻击大型猎物,消耗猎物的体力。

异特龙与剑龙的骨架 - 丹佛自然历史博物馆
埃米莉·雷菲尔德(Emily J. Rayfield)等人使用有限元分析,研究了异特龙的头颅骨,也得到了类似的结果。根据其中的生物力学研究结果,异特龙的头颅骨非常强壮,但咬合力相当小。异特龙的咬合力只有805到2,148N,少于短吻鳄(13,000 N)、狮子(4,167 N)、美洲豹(2,168 N),原因是它们咬合时只使用到颌部的肌肉,然而它们的头颅可承受约55,000牛顿来自于齿列的垂直压强。这个研究也提出异特龙使用头部来撞击猎物,并张开大口、撕咬猎物,而不用撑开头部的骨头。这个研究认为异特龙的头部结构允许它们采取不同的猎食模式来攻击不同的猎物:它们的头部较轻型,可攻击较小、较灵活的鸟脚类恐龙;但头部有足够的强度承受撞击,可允取它们采取伏击方式攻击较大型的剑龙科与蜥脚类恐龙。其他的科学家则对这个研究表示异议,他们认为现存生物中没有采取撞击方式猎食的动物,并提出异特龙的头部有足够的强度,应可承受猎物挣扎的力量。雷菲尔德等人对异议提出回应,他们承认现存生物中没有类似异特龙的动物,但异特龙的齿列适合这种攻击方式,而它们的头部结构可保护上颌、减低承受力量。另一种可能则是,兽脚类恐龙(例如异特龙)不必费力将猎物杀死,而是从活生的蜥脚类恐龙身上咬下足够的肉块,肉块的大小只需维持猎食动物生存即可。这种猎食方式也使猎物有机会痊愈,而猎食者也可能以类似的方式再度猎食。另外,鸟脚类恐龙是当地最常见的猎物,因此异特龙可能采偷袭方式猎食鸟脚类恐龙,使用前肢抓住猎物,并咬断猎物喉咙的气管,类似今日的大型猫科动物。异特龙的前肢强壮,能够抓紧猎物,所以这个猎食方式是可能成立的。
其他影响进食方式的因素包含:眼睛、前肢、以及后肢。异特龙的头部形状将立体视觉限制在20°的范围内,略小于现代鳄鱼。如同鳄鱼,这个范围已足够异特龙判断猎物的距离与攻击时机。相较于其他兽脚类恐龙,异特龙的前肢适合抓住一定距离的猎物,还有将猎物拉近;而指爪的构造显示它们可用来勾取物体。经估计,异特龙的最高奔跑速度可达每小时30到55公里。
社会行为
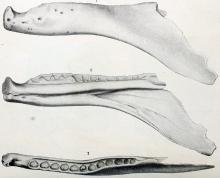
Labrosaurus ferox的正模标本齿骨,伤痕可能是由其他异特龙所造成
自从1970年代以来,半科学文献与大众读物都将异特龙描述成以群体方式猎食,并以蜥脚类恐龙与其他大型恐龙为猎食对象。罗伯特·巴克(Robert T. Bakker)从脱落的牙齿、大型猎物的被咬过骨头研判,异特龙具有亲代养育的社会行为。巴克认为成年异特龙将食物带到巢穴中,以供幼年异特龙食用,并防止其他肉食性动物找到它们的食物。但实际上,很少证据显示兽脚类恐龙具有群居行为。而腹肋上的伤口、头颅上的咬痕,则证明异特龙具有互相攻击的物种内行为;Labrosaurus ferox的下颌被怀疑有病状,有可能是互相攻击的后果。同一群体内的异特龙,或两只敌对的个体,可能借由这种互相咬伤头部的行为,来确定活动范围。
早期的研究认为异特龙可能会采群体方式猎食。但近年的研究提出异特龙与其他兽脚类恐龙具有侵略性的物种内行为,而非合作性的行为,如同其他的双孔动物。一个研究则推论兽脚类恐龙会合作猎食,而非个别猎食;这种行为在脊椎动物中较少见,而现存双孔动物(包括蜥蜴、鳄鱼、鸟类)很少合作猎食。许多现代的掠食性双孔动物是领域性,会将侵入领地的同类杀死、并吞食它们的尸体;另外当聚集在食物周围时,它们会将企图抢先的较小个体杀死。克利夫兰劳埃德采石场的大量异特龙化石可能导因于它们在同类相食时淹死。这也可以解释异特龙化石中,幼年与亚成年个体所占的比例较高;因为在现代的鳄鱼与科莫多龙中,幼年与亚成年个体较少在聚食地点中被杀死。这理论也可解释巴克所发现的巢穴状况。有些证据显示异特龙具有同类相食的行为,例如肋骨碎片上有脱落的同类牙齿、一个肩胛骨上有可能的齿痕、巴克所发现的巢穴中的异特龙骨骸可能遭到同类相食。
脑部与感觉
一个针对异特龙脑部的电脑断层扫描,发现它们的脑部与鳄鱼有较多的共同点,而与现代鸟类脑部的共同点较少。前庭器官的结构显示它们的头部保持在几乎水平的位置,而非朝上或朝下。内耳的结构类似鳄鱼,所以异特龙可能容易听到低频的声音,但难以听到细微的声音。异特龙的嗅球大,可能适合感觉气味,但辨别气味的区域相当小。
大众文化

异特龙的模型,位于波兰Bałtów
在大众文化中,异特龙与暴龙皆是大型肉食性恐龙的代表。异特龙也是博物馆常见的恐龙之一,部分原因为克利夫兰劳埃德恐龙采石场所出土的大量异特龙化石;到了1976年,已经有三大洲、八个国家的38个博物馆具有从克利夫兰劳埃德恐龙采石场所出土的异特龙化石。异特龙是犹他州的官方州恐龙。
自20世纪初期开始,异特龙便已出现在大众文化之中。在阿瑟·柯南·道尔(Arthur Conan Doyle)的小说《失落的世界》(The Lost World)以及1925年的电影版本中,异特龙被描述成顶级掠食动物;暴龙同时也出现在电影版本中。异特龙也出现在两部结合恐龙与西部片的电影中:《荒山大贼王》(The Beast of Hollow Mountain, 1956)、《暴龙关吉》(The Valley of Gwangi, 1969)。在《暴龙关吉》中,著名电影特效师雷·哈利豪森(Ray Harryhausen)根据暴龙的形象创造出关吉,但关吉却被标名为异特龙。哈利豪森常将两者搞混,他在DVD的访谈中透露:“它们都是肉食性动物,看起来都很凶暴…,只是其中一个稍微大于另外一个”。
异特龙也出现在BBC的电视节目《与恐龙共舞》(Walking with Dinosaurs)的第二集与第五集。而《与恐龙共舞》的特别节目《异特龙之谜》(Ballad of Big Al),则是以著名的“大艾尔”作为主角,叙述了它的一生。异特龙也出现在迪士尼频道制作的电视节目《恐龙王朝》(Dinosaur Revolution)。
在CAPCOM于2000年推出的电子游戏《恐龙危机2》中,异特龙也有多次登场。
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

相关资料





- 有价值
- 一般般
- 没价值















推荐阅读




关于我们

APP下载


