






驰龙科
叙述
身体架构

伶盗龙的骨架,位于布鲁塞尔
驰龙科的独特身体架构,有助于恐龙是种活跃且快速动物的理论,以及恐龙与鸟类是近亲的理论。在罗伯特·巴克(Robert T. Bakker)替约翰·奥斯特伦姆(John Ostrom)的1969年专题论文所做的图解中,将恐爪龙描绘成快速奔跑的姿态,这成为史上最具影响力的古生物学重建图。驰龙科具有大型头部、锯齿边缘牙齿、狭窄口鼻部、眼睛向前,显示具有一定程度的立体视觉。如同大部分的兽脚类恐龙,驰龙科的颈部长,呈S状弯曲,身体相当短。它们的手臂长,某些物种的手臂可以在胸前交叉,手部相当大,上有三根长手指,第一指最短,第二指最长,手指上有大型指爪。耻骨末端大,往尾巴基部延伸。第二脚趾上有大型、弯曲趾爪。尾巴修长,尾椎长而低平,第14节尾椎之后缺乏横突(Transverse process)与神经棘。
目前已发现部分驰龙科的身体覆盖者羽毛,包含前肢与尾巴上的大型、片状羽毛,可能所有的驰龙科都有羽毛。这个理论是在80年代中晚期首次出现,并在1999年得到证实。这些改变导致驰龙科在电影与重制模型中的形象不断改变。
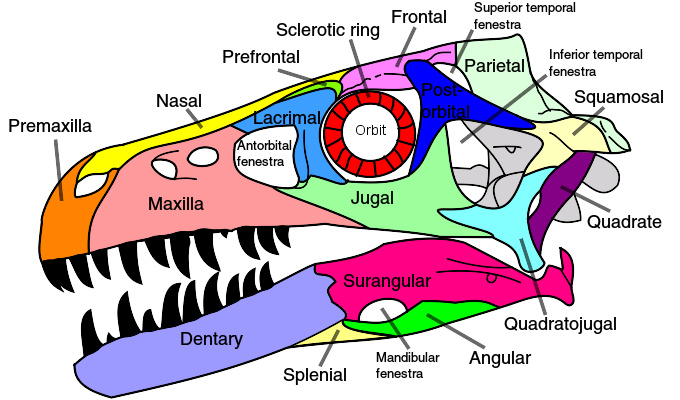
驰龙的颅骨构造图
鉴定特征
驰龙科具有以下特征:额骨短而呈T字形,构成上颞孔的前缘、方骨的后突接触方轭骨、背椎的椎体横突对(Parapophyses)竖起、独特的第二脚趾、人字形骨与尾椎的前关节突延长,延伸至前几节脊椎、鸟喙骨上有关节盂孔。
体型
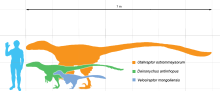
犹他盗龙(黄)、恐爪龙(绿)、伶盗龙(蓝)与人类的体型比较图
驰龙科是群小型到中型的恐龙,身长范围从0.7米的大黑天神龙,到超过6米的犹他盗龙与阿基里斯龙。杨百翰大学的某些犹他盗龙标本,身长可能达约11米。但这些化石还没有经过详细研究,经过详细研究后可能会产生不同数值。驰龙科可能两次演化出大型体型,一次是驰龙亚科的犹他盗龙、阿基里斯龙,另一次是半鸟亚科的南方盗龙,其身长估计约5米。在英格兰威特岛发现的个别大型牙齿,可能代表某种大型驰龙科恐龙,体形接近于犹他盗龙。根据牙齿形状,它们可能属于伶盗龙亚科。如果属实,可能代表驰龙科分别三次演化出大型体型。
目前所发现最原始的驰龙科是大黑天神龙,身长只有70公分,也是目前已知最小的驰龙科恐龙。大黑天神龙、小盗龙的发现,加上伤齿龙科的近鸟龙,显示近鸟类的共同祖先可能非常的小,身长约65公分,体重约600到700公克;近鸟类包含驰龙科、伤龙科、以及鸟类。
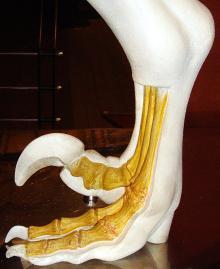
驰龙科的脚部构造模型
脚部
如同其他兽脚类恐龙,驰龙科以后肢行走。然而,大部分兽脚类恐龙以后肢的三根脚趾接触地面;而驰龙科只有用第三、第四脚趾支撑身体重量,属于双趾型动物。从足迹化石显示,它们的第二趾明显地后缩。第二脚趾上有大型、镰刀状、弯曲趾爪,行走时会往后缩起,被认为是作为杀死猎物使用,或是攀爬树木。真驰龙类的体型较大,第二趾爪的特化程度更为明显。真驰龙类的其中一属,巴拉乌尔龙(Balaur)的第一趾、第二趾都朝向脚掌前方,这两趾都向上后缩,都具有大型、镰刀状趾爪。
尾巴
驰龙科具有长尾巴。大部分的尾椎有骨质、棒状前关节突,某些物种的尾巴则具有骨化肌腱。在奥斯特伦姆的恐爪龙研究中,他提出这些特征将尾巴硬挺,不易弯曲,惟有尾巴基部可灵活活动。但是,蒙古伶盗龙的一个保存良好标本(编号IGM 100/986),尾巴向后弯曲呈S状,关节没有脱落,这显示它们的尾巴可做出一定程度的平行运动。驰龙科的尾巴被假设在奔跑或滑翔时,作为稳定器或平衡器使用;在顾氏小盗龙与中国鸟龙的一个标本(编号 NGMC 91)的尾巴末端,有羽毛构成的小型、钻石形扇状物,可能作为气动性上的稳定器,以及方向舵使用。
羽毛

顾氏小盗龙的羽毛痕迹
有许多的证据显示,驰龙科的身体覆盖者羽毛。有些驰龙科化石的手部、手臂、尾巴,保存者长的正羽,而身体则保存者绒羽。另外有些化石,虽然没有保存羽毛,但前臂骨头上则有突起处,可能在生前是羽毛的附着处。整体而言,驰龙科的羽毛型态非常类似始祖鸟。
第一种被发现有明显羽毛证据的驰龙科恐龙是中国鸟龙,由徐星等人于1999年在中国发现。许多其他的驰龙科化石曾发现羽毛覆盖者身体,有些拥有完全发展的有羽毛翅膀。有些物种甚至显示出后肢为第二对翼的证据,包括小盗龙与羽龙。因为羽毛痕迹只会在细致的沉积层中保存下来,有些发现于粗糙岩层中的化石保有了羽茎瘤,羽茎瘤是鸟类的飞羽附着点。胁空鸟龙与伶盗龙的化石都发现了羽茎瘤,显示它们的前肢具有羽毛。因此,大型、居住于地面的驰龙科恐龙极可能拥有羽毛,如同大部分的现代无法飞行鸟类仍保有它们的飞羽。虽然有些科学家主张,大型驰龙科失去了部分或全部的羽毛,但伶盗龙的羽茎瘤被视为是所有的驰龙科都具有羽毛的证据。
与鸟类的关系
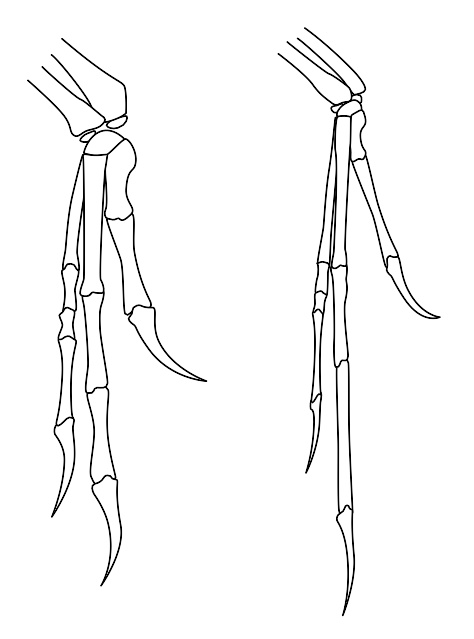
恐爪龙的手(左)与始祖鸟的手(右)比较。由此可看出驰龙科与鸟类的相似处
驰龙科属于兽脚亚目,与早期的鸟类有许多共同特征。有许多研究讨论过驰龙科与鸟类的关系,随者许多新证据的出现,驰龙科与鸟类关系的假设也不断改变。在2001年,马克·诺瑞尔(Mark Norell)与其同事研究大量的虚骨龙类化石,并提出驰龙科是鸟类的最近亲,两者与伤齿龙科共同属于鸟翼类。诺瑞尔更指出,驰龙科是鸟翼类中的一个并系群。在2002年,S.H. Hwang与其同事参考了诺瑞尔的研究与新发现化石,提出鸟类是驰龙科、伤齿龙科两者的近亲。
古生物学界目前的共识多同意Hwang等人的研究,驰龙科与伤齿龙科是最近亲,两者共同组成恐爪龙下目。而恐爪龙下目是鸟类的姊妹分类单元,也是鸟类的最近亲。但古生物学界还没有足够证据可指出驰龙科(以及它们的祖先)能否飞行或滑翔。
相关理论
驰龙科传统上被认为是非鸟类的恐龙,然而有些研究人员认为驰龙科也属于鸟纲演化支。首先,驰龙科具有羽毛,符合传统的鸟类定义-具有羽毛。羽龙意为“隐藏的飞行者”,它们最初因其具有羽毛而被命名为鸟类,造成许多误解。
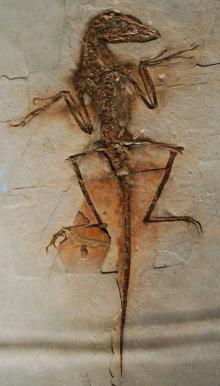
中国鸟龙的可能标本
目前至少两派研究人员提出驰龙科是从可飞行的祖先演化而来。其中一派主张鸟类先演化出现,乔治·奥利舍夫斯基(George Olshevsky)被认为是这理论的首位提出者。葛瑞格利·保罗(Gregory S. Paul)则提出许多证据,指出驰龙科演化自会飞行的恐龙,可能类似始祖鸟。根据此理论,体型较大的驰龙科是在演化过程中失去飞行能力,如同新西兰的已灭绝恐鸟、以及鸵鸟。在1988年,保罗提出驰龙科与现代鸟类的关系,比始祖鸟与现代鸟类的关系更为接近。但在2002年,保罗改提出驰龙科是始祖鸟的最近亲。
在2002年,S.H. Hwang与其同事提出小盗龙是最原始的驰龙科。在2003年,徐星与其同事根据小盗龙的羽毛、翅膀等特征,提出驰龙科的祖先能够滑翔。而体型较大的驰龙科则在演化过程中失去滑翔能力,成为陆栖动物。
在2002年,斯特芬·柯瑞克斯(Stephen Czerkas)叙述、命名羽龙。柯瑞克斯错误地判断羽龙的羽毛只位在前肢与手部,将羽龙重建成双翼恐龙;并认为羽龙较接近鸟类,而非滑翔动物。但是,日后有其他科学家提出羽龙可能是小盗龙的次异名。而柯瑞克斯也在他的著作中更正他的错误。
另一派研究人员,例如赖利·马丁(Larry Martin),则提出驰龙科与所有手盗龙类都不是恐龙。马丁并提出鸟类与手盗龙类并非近亲,但在2004年,马丁修正他的理论,认为手盗龙类是鸟类的近亲。但马丁认为手盗龙类在演化过程中失去飞行能力,而鸟类演化自非恐龙的主龙类,而兽脚亚目的大部分成员并不属于恐龙。

瑟马普利斯始祖鸟标本,具有类似驰龙科的第二趾爪。
在2005年,Mayr与Peters叙述了一个在怀俄明州瑟莫普利斯镇发现的良好始祖鸟标本,发现始祖鸟的生理结构比原先认知的更类似恐龙。另外,他们还具有类似驰龙科的大幅弯曲第二趾爪、原始上颌、未反转后趾。Mayr与Peters的种系发生学研究提出孔子鸟与小盗龙的亲缘关系更为接近,而离始祖鸟较远,使鸟翼类成为并系群演化支。他们更提出其近鸟类祖先能够飞行或滑翔,而驰龙科与伤齿龙科在演化过程失去飞行或滑翔能力。Corfe与Butler则以研究方法层面提出反对意见。
在2007年,另有其他的理论出现。艾伦·特纳(Alan Turner)与其同事命名了一种新驰龙科-大黑天神龙,大黑天神龙是最基础、原始的驰龙科,比小盗龙更原始。大黑天神龙具有短手臂,无法滑翔。特纳等人认为飞行能力演化自鸟翼类,而驰龙科的祖先无法飞行、滑翔。根据亲缘分支分类法研究,原始的大黑天神龙显示驰龙科的祖先无法飞行。
古生物学

恐爪龙的骨骼

驰龙的脚部与第二趾爪
指爪与第二趾爪的功能
关于驰龙科第二脚趾的镰刀状趾爪的功能,目前存在着不同的意见。在1969年,当约翰·奥斯特伦姆(John Ostrom)叙述恐爪龙的镰刀状趾爪时,他将这些趾爪解释为类似刀片的砍伤用武器,相当类似剑齿虎的犬齿,恐爪龙使用强壮的趾爪来挖开猎物的身体。在1987年,另有研究认为它们会以第二趾爪猎杀大型角龙类恐龙。这些解释被应用到所有驰龙科恐龙身上。然而,菲利普·曼宁(Phillip Manning)等人认为这些趾爪并非当做钩爪使用,并将它们重建为长椭圆形横断面,外围包覆者角质层,而非原先认为的倒泪滴形横断面。在曼宁的解释中,第二趾爪可能在猎食大型猎食时,用来协助攀爬用,同时也是种刺伤武器。
在1970年,约翰·奥斯特伦姆曾参考鸵鸟与食火鸡,发现这些鸟类会用较大型的第二趾爪攻击猎物,进而产生恐爪龙会利用第二趾爪攻击猎物的理论。食火鸡的第二趾爪可长达12.5公分。奥斯特伦姆并引用一个1958年的鸟类论文,认为后肢与第二趾爪,可用来将猎物的身体撕开、将内脏挖出。在近年,Chhristopher P. Kofron研究241件食火鸡的攻击纪录,发现总计有1个人类、2只狗的被攻击死亡案例,但没有证据显示食火鸡会用第二趾爪将猎物的内脏挖出。相反地,食火鸡将趾爪作为防卫武器,攻击对它们产生威胁的动物、或威吓对方叫鹤也具有较大的第二趾爪,却是用在撕裂小型猎物,以适合吞咽。
在2009年,菲利普·曼宁等人着手研究第二趾爪与前肢指爪的功能。他们从生物力学方面开始研究,研究第二趾爪与前肢指爪在受力时,压力与张力的分布与传递状况。他们采用X光线成像技术,将一个伶盗龙的前肢指爪建立出3D立体图像。接者,他们将雕鸮的指爪作为对照组。借由比对发现,伶盗龙的前肢指爪相当适合用来攀爬。其尖锐的指爪尖端,适合穿刺、固定;而弯而宽的指爪基部,有助于平均地传递受力时的压强。
曼宁等人同时也研究第二趾爪的弯曲程度,并以现代鸟类与哺乳类作为对照组。根据早期的研究,趾爪的弯曲程度,与该动物的生活方式息息相关。趾爪大幅弯曲的动物,通常擅于攀爬;趾爪较笔直的动物,通常是在地面生存、活动的动物。曼宁等人发现,恐爪龙的第二趾爪,弯曲程度可达160度,属于攀爬动物的范围。同样地,驰龙类的前肢指爪,弯曲程度也属于攀爬动物的范围。
对于曼宁等人的研究,彼得·马克维奇(Peter Mackovicky)有不同的看法。他认为小而原始的驰龙类(例如小盗龙)可能是攀爬动物,但晚期的大型驰龙类(例如阿基里斯龙)虽然具有大、弯曲的第二趾爪,但体型过大,已无法攀爬树木。马克维奇推测,晚期的大型驰龙类的第二趾爪,已失去攀爬功能,但会在猎食时,用来攻击猎物。
在2009年,菲力·森特(Phil Senter)发表一份驰龙科脚趾研究,提出部分物种可能会使用后肢趾爪挖开昆虫的巢穴。某些小型驰龙科,例如:胁空鸟龙、鹫龙,可能有时会以昆虫为食;而体型较大的驰龙科,例如:恐爪龙、内乌肯盗龙,也可能使用后肢趾爪挖开地表洞穴,以里面的小型脊椎动物为食。但菲力·森特并没有测试大幅弯曲的第二趾爪,在挖掘洞穴时是否有发生功能。
在2011年的一项恐爪龙类趾爪研究,也认为第二趾爪无法作出有效的刺穿、划开猎物身体的动作,并提出第二趾爪有固定猎物功能的理论,认为驰龙科可能以小型猎物为食。这些研究比对不同恐爪龙类、现代鸟类的脚掌结构、趾爪形状。研究发现,许多掠食性现代鸟类也有大型第二趾爪,当它们猎食小型猎物时,会使用脚掌、趾爪将猎物固定在地面上,用体重施压于猎物,再用鸟喙吞食这些猎物,尤其是鹰科。不只是掠食性鸟类,其他掠食动物猎食时,也会用趾爪紧握、固定小型猎物。研究发现恐爪龙的脚掌、脚趾应有大型肌腱,跖骨短,第一趾(后趾)偏向后方,这些特征很符合固定猎物功能的特征。研究推测,当恐爪龙等进阶型物种猎食时,可能会用第二趾爪攻击、固定猎物到地面上,然后头低下撕咬、吞食猎物,而尾巴具有平衡身体的功能。它们的牙齿后侧才有大型、钩状边缘,比较适合拔出猎物的肉体,而非撕裂。前肢强壮,有大型指爪,但覆盖者长羽毛,当驰龙科猎食时,可能会将前肢张开,以伺机保持身体的平衡。以上特征也支持固定猎物功能的理论。
某些原始驰龙科、以及伤齿龙科的第二趾爪,还没有演化成后期恐爪龙类的大型、镰刀状趾爪。它们的脚趾关节允许作出更灵活的动作,而后期恐爪龙类的第二脚趾趾能作出向上后缩、向下的动作。这有可能是它们以更小型动物为食,因此不用演化出大型趾爪、强壮脚掌以固定猎物。研究人员推测,近鸟类可能共同拥有这些演化特征,而现代鸟类则继承这些特征。

打斗中的原角龙与伶盗龙
群体猎食
曾经在腱龙的化石附近发现一个小群体的恐爪龙化石;腱龙是种大型草食性鸟脚下目恐龙。这发现被解释成驰龙科恐龙以有秩序群体猎食的证据,类似某些现代哺乳类。然而,并非所有古生物学家认为这证据具决定性,而后来的研究显示恐爪龙较可能以无秩序的群体攻击猎物。现代鸟类与鳄鱼(驰龙科的近亲们)显示出很少协调的猎食行为;相反地,它们通常是独自猎食,或是被尸体所吸引,同一物种之内可能存在者这两种不同的方式。以印尼的科摩多龙来说,它们在群体进食的时候,最大型的个体会先进食,并攻击企图先进食的较小型个体;如果较小的个体死亡,将会遭到同类相食。当以这个现象来观察原本被认为是群体猎食的驰龙科恐龙时,则呈现出同时符合科摩多龙与鳄鱼的进食模式。这些地点所发现的恐爪龙骨骸都是来自于未成年体,而遗失的部分可能被其他恐爪龙所吞食,这不符合原先所认为的群体猎食行为。
在2007年,科学家们在中国山东省发现了第一个大量的驰龙科足迹化石。这个足迹化石可能是由体型接近阿基里斯龙的大型驰龙科恐龙所留下,而且是由六个相近大小的个体,沿者海岸共同前进。这些个体相距约一米,朝同一方向缓慢前进。研究人员提出,这个足迹化石可证明驰龙科的某些物种过者群体生活。虽然该足迹化石并不能明确指出驰龙科的猎食行为,但无法排除它们以群体猎食的可能性。此外,该发现还支持了驰龙科在行走时,第二脚趾后缩、离地的假设。

奥氏胁空鸟龙,曾一度被归类于鸟类

以四翼方式滑翔飞行的顾氏小盗龙
飞行与滑翔能力
目前至少有两种驰龙科恐龙被认为具有飞行或滑翔能力,胁空鸟龙、顾氏小盗龙。胁空鸟龙最初被归类于鸟类,后来的研究将其归类于驰龙科。胁空鸟龙的长前肢被发现有羽茎瘤,羽茎瘤是飞羽的附着点,因此胁空鸟龙被认为可以动力飞行。从结构来看,胁空鸟龙的前肢比始祖鸟更有力,并附有强壮的韧带,可以作出拍动的动作,展开飞行。路易斯·齐亚比(Luis Chiappe)提出胁空鸟龙具有这些适应特征,可以飞行,但飞行能力较现代鸟类差。
顾氏小盗龙的前肢与后肢相当发达,科学家认为它们可以采四翼方式滑翔。在2005年,萨克·查特吉(Sankar Chatterjee)提出顾氏小盗龙的四肢类似双翼飞机,但是采前后排列方式。查特吉认为它们在树枝向下俯冲,以U字形的曲线滑翔,然后降落在地表或是其他的树上;尾巴与后肢可用来控制方向与速度。此外,查特吉认为顾氏小盗龙有初步的飞行能力。
羽龙也被认为可能具有飞行或滑翔能力,但有科学家提出羽龙可能是小盗龙的异名。
感应器官
在2011年,科学家比较小盗龙、中国鸟龙、伶盗龙、现代鸟类与爬行动物的巩膜环大小,提出小盗龙、中国鸟龙可能属于夜行性动物,而伶盗龙属于无定时活跃性的动物,觅食、移动行为跟白天黑夜没有正相关,只休息短暂时间。近年研究发现,小盗龙具有色彩斑驳的羽毛颜色,由于羽毛颜色鲜艳的现代鸟类,都是日行性动物,显示小盗龙并不是夜行性动物。
驰龙科的嗅球/身体比例,接近其他兽脚类恐龙、现代鸟类,例如暴龙科与火鸡秃鹰,显示驰龙科具有良好的嗅觉,有助于它们寻找猎物。
病征
在2001年,布鲁斯·罗斯柴尔德(Bruce Rothschild)等人发表一份兽脚类恐龙的压力性骨折、肌腱撕裂伤研究。他们发现驰龙科的压力性骨折迹象,大多是多次事件造成的伤口,较少来自于单一事件,因此推论它们的压力性骨折较常导因于习惯性动作,较少来自于外力。他们在两个手掌指爪发现扭伤骨折的迹象,其中一个应是由压力性骨折造成的。脚部的压力性骨折,可能导因于奔跑、长期移动;手部的压力性骨折,则可能导因于捕抓猎物时,猎物的全力挣脱。
分类系统
分类学
在1992年,W.D. Matthew与巴纳姆·布郎(Barnum Brown)建立了驰龙亚科,当时只包含驰龙属,属于现在已废止的恐爪龙科。目前,驰龙科与伤齿龙科共同组成恐爪龙下目。
驰龙科的各亚科经常因为新的研究而改变内容,但大致上包含以下各演化支。而且许多属的化石保存状态不好,无法分类到任一亚科之中,例如最原始的大黑天神龙。
半鸟亚科(Unenlagiinae)是驰龙科中最原始的亚科。半鸟亚科是个充满谜题的演化支,是最少得到理论支持的亚科,其中有些或全部物种可能并不属于驰龙科。其中较大、生活于地面的物种,例如鹫龙与半鸟,显示出强烈的飞行适应特征,但它们可能过大而不能飞行。另一个物种胁空鸟龙的体型非常小,拥有发展良好的翼,并有羽茎瘤(飞羽的附着点)的证据,它们即有可能有飞行能力。
小盗龙亚科(Microraptorinae)是驰龙科中第二原始的演化支。小盗龙亚科包含许多最小的驰龙科恐龙,并显示出树栖生活的证据。所有驰龙科的皮肤痕迹证据都来自于小盗龙亚科,所有小盗龙亚科都覆盖者羽毛并拥有发展良好的翼。如同半鸟亚科,有些物种可能拥有主动飞行能力。
伶盗龙亚科(Velociraptorinae)传统上包含伶盗龙、恐爪龙,而新发现的白魔龙可能属于伶盗龙亚科,而蜥鸟盗龙的分类位置仍未确定。
驰龙亚科(Dromaeosaurinae)通常包含体型中到大的物种,并拥有箱形的头颅骨;其他的亚科通常拥有狭窄的口鼻部。
蜥鸟盗龙亚科(Saurornitholestinae)是尼克拉斯·朗里奇(Nicholas Longrich)与菲力·柯尔(Phil Currie)在2009年建立的新分类单元。西爪龙的发现,使得斑比盗龙不属于小盗龙亚科。于是他们建立蜥鸟盗龙亚科,以包含蜥鸟盗龙与近亲斑比盗龙。

大黑天神龙,一种原始驰龙科

西爪龙,属于小盗龙亚科

半鸟,属于半鸟亚科

犹他盗龙,一种大型的驰龙亚科

斑比盗龙,属于蜥鸟盗龙亚科

伶盗龙,属于伶盗龙亚科
以下驰龙科各属的分类表是根据赛里诺(2005年)、森特(2004年)、马克维奇等人(2005)、诺瑞尔等人(2006年)、特纳(2007年)等人、以及汤玛斯·荷兹(2011年)的研究而来。
驰龙科 Dromaeosauridae
系统发生学
在1998年,保罗·塞里诺(Paul Sereno)将驰龙科定义为一个演化支:包含驰龙,但不包含伤齿龙、似鸟龙、或家麻雀在内的最大天然族群演化支。驰龙科的不同亚科都曾被定义为演化支过,都常是以驰龙或其他次演化支作为定义基准点;举例而言,马克维奇将半鸟亚科定义为:在驰龙科中,所有亲缘关系接近半鸟,而离伶盗龙较远的所有物种。小盗龙类(Microraptoria)是唯一不是亚科的次演化支,森特与其同事在建立时,没有加上亚科的字尾-inae,以避免被误认,并独立于驰龙科之外。但塞里诺则提出不同的定义,并将小盗龙类改名为小盗龙亚科(Microraptorinae),归类于驰龙科之内。但是,这个用法仅出现在他的TaxonSearch database网站,且未在正式文献现过。
以下演化树是根据尼克拉斯·隆瑞奇与菲力·柯尔在2009年的研究,其中的亚科是采用赛里诺在2005年的定义。
以下演化树来自于菲力·森特等人的2012年研究:
以下演化树来自于艾伦·透纳等人的2012年研究:
备注:有些研究人员以及亲缘分支分类法研究,尤其是针对驰龙科中的小型可飞行物种的研究,发现始祖鸟是驰龙科的早期成员。如果始祖鸟属于驰龙科,根据国际动物命名委员会的规定,始祖鸟科将比驰龙科还具有优先权。因此若将始祖鸟包含在内,那驰龙科将改名为始祖鸟科。保罗·塞里诺是首批采用始祖鸟科,而不采用驰龙科的科学家之一。
大众文化

覆盖羽毛版的驰龙模型
1993年的电影《侏罗纪公园》(Jurassic Park)中所出现的伶盗龙,体型比该属所发现最大的标本还大。实际上,这些“迅猛龙”是其近亲恐爪龙。 葛瑞格利·保罗(Gregory S. Paul)在他的书籍《Predatory Dinosaurs of the World》中,将恐爪龙列为伶盗龙的一种,并将该种命名为平衡伶盗龙(Velociraptor antirrhopus),这个分类法后来遭到普遍地否决。麦克·克莱顿(Michael Crichton)将这个观点写入他的《侏罗纪公园》系列小说当中,采用葛瑞格利·保罗的说法,并被应用到电影当中。在电影《侏罗纪公园》现的驰龙科恐龙,在当时是正确的描述,但现在看来则有许多方面是错误的,包含缺乏羽毛。但在《侏罗纪公园 III》(Jurassic Park III)中则将这个错误纠正回来。
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

相关资料





- 有价值
- 一般般
- 没价值















推荐阅读


关于我们

APP下载


