





-
 QQ空间
QQ空间
-
 QQ好友
QQ好友
-
 微信好友
微信好友
-
 新浪微博
新浪微博
进化
名词起源
英语
英文中的“evolution”一词,起源于拉丁文的“evolvere”,原本的意思是将一个卷在一起的东西打开,当今的解释是在地球历史上,不同种类的生物怎样从早期的形式发展出来的一个过程。当代英语字典也有“发展”、“从经历中获取”之义。
在1859年出版的《物种起源》第一版至1876年的第六版,达尔文也以“evolved”这个字结尾。当时达尔文是使用“经过改变的继承”(descent with modification)、“改变过程”(process of modification)或是“物种改变的原理”(doctrine of the modification of species)等。他曾在《物种起源》第7章中说,天择的最后结果,包括了生物体的进步(advance)及退步(retrogression)两种现象”,而总括来说,从地球有生物开始,是进步或退步的问题放在第10章讨论,结论是现有的生物都是经过长时间的进步,到某程度而没有违反天择的,发展便停留下来。
在《物种起源》最后第六版本,达尔文用了“Evolution”这个字多次,还很直接明显地称他的理论为“Theory of evolution”。达尔文在结语中说:
不同的翻译
物种起源的第一本中文版由马君武翻译,与较早期的日语版本同用现在通行的“进化”一词,有学者认为来源是和制汉语。早于达尔文逝世不久,中文上用“进化论”是一致的。
进化思想史

卢克莱修1842年,查尔斯·达尔文写出了物种起源的第一份草稿。阿尔弗雷德·拉塞尔·华莱士
早在古希腊时代,类似进化的思想已经出现,例如前苏格拉底哲学家阿那克西曼德及恩培多克勒认为一种生物可以源于另一种生物。这种思想一直延续到罗马时代,继恩培多克勒之后,卢克莱修在他的作品《物性论》中也提及类似进化的思想。相对于这些唯物主义的思想,亚里士多德了解不仅是活的东西,对一切自然的东西而言,进化是对不同的自然可能性的一种不完美实现,如理型、理念、物种。这是他理解大自然目的论的一部分:一切事物都在神圣的(英语:Divinity)宇宙秩序中发挥预期作用。在中世纪,这种思想的变体成为标准理解和被纳入基督教的知识,但亚里士多德没有要求生物体总是对应形而上一对一的形式和没有就新生物的产生给出具体例子 。
在17世纪,现代科学的新方法反对亚里士多德的说法,它要求对自然现象的解释在对所有看得见的事物上,其遵从的物理定律要相同,但这并没有要求任何固定的自然属类或神圣的宇宙秩序需要存在。然而,这种新方法是在生物科学上缓慢扎根。约翰·雷,首次把更概括的术语“种”(species)应用于植物和动物种类上,但他严格地标识每类活着的东西为一种物种,建议每一个物种可以由一代延续一代的自身特点来定义。他认为这些物种是上帝设计的,但表现出局部条件的差异。在卡尔·林奈于1735年引入的生物分类中,其明确承认种关系的分层特性,但根据“上帝的计划”物种仍被视为不变。
同期的其他博物学家根据自然规律,推断出物种会随时间演变。1751年,皮埃尔·莫佩尔蒂写道透过繁殖和多代累积的自然变更,会产生新的物种。乔治-路易·勒克莱尔,布丰伯爵认为物种可以蜕化成不同的生物,伊拉斯谟斯·达尔文则提出所有恒温动物可能是单一微生物的后裔。1809年,拉马克提出第一个全面的进化机制,其猜想自然发生在具有固定的渐进趋势的平行世系中不断地把简单的生命变为更复杂的生命,并推测在局部的层面,下一代会通过遗传他们父母用进废退的器官以适应环境。(后一方法后来被称为拉马克主义。)这些想法受博物学家谴责,因为这些推测缺乏实证支持。尤其乔治·居维叶坚持认为种是不相关的并不变,它们的相似只是反映了上帝为了其功能需求的设计。英国牧师威廉·佩利(英语:William Paley)把林奈的思想在《自然神学》(英语:Natural Theology or Evidences of the Existence and Attributes of the Deity)中进一步发展,其中提出复杂的适应度是上帝所设计,这亦是查尔斯·达尔文推崇的证据。
自然选择理论是一个关键理念把不变类型学或生物类型的概念打破,这是达尔文部分受到马尔萨斯的《人口论》影响而建立的理论,达尔文指出,人口增长会导致“生存斗争”,有利的变化占了上风,其他的则死去。每代中,很多子女因有限的资源而在达至生殖年龄前无法生存,这可以解释共同祖先透过自然法则以同样方式对待所有有机体进化而来的植物和动物的多样性。从1838年起,达尔文逐步建立了他的“自然选择”理论,在1858年,当华莱士给他发了一封理论几乎相同的的信时,他正在编写关于这个议题的“著作”。他们俩的共同论文“论物种形成变种的倾向;以及论这些变种和物种经由自然选择之手段而得以延续的方式(英语:On the Tendency of Species to form Varieties; and on the Perpetuation of Varieties and Species by Natural Means of Selection)”在1858年的伦敦林奈学会上发表。在1859年年底,达尔文关于其“理论”的出版物《物种起源》详细解释自然选择及其机制,造成日后渐为人接受的进化概念。托马斯·亨利·赫胥黎把达尔文的思想应用至人类,并利用古生物学和比较解剖学,去证明人类和类人猿有着共同的祖先。有些人对这种思想感到不安,因为它暗示人类在宇宙中并没有特殊的地位。
生殖遗传和新性状起源的确切机制仍然是个谜,为此达尔文曾提出他的临时学说泛生论。1865年,孟德尔遗传性状是在可预测的方式,通过元素的自由组合(孟德尔第二法则)和分离遗传(该元素后来被称为基因),孟德尔遗传定律最终取代大部分达尔文的泛生学说。奥古斯特·魏斯曼提出产生可产生配子的生殖细胞和身体的体细胞之间的重要区别,这表明遗传只经生殖细胞系(英语:Germline)发生。许霍·德弗里斯把达尔文的泛生论连接至魏斯曼的发现并提出达尔文的泛生论应集中于细胞核,他表示细胞核可以移动到细胞质去改变细胞结构。德弗里斯也是令孟德尔的研究知名的研究人员之一,认为孟德尔性状对应于沿种系的遗传变异传递。为了解释新变种如何起源,德弗里斯制定了一个突变理论,导致与德弗里斯同阵的生物统计学派(Biometricians)及达尔文的进化论出现暂时性裂痕。20世纪30年代,群体遗传学领域的先驱如罗纳德·费雪、生物学家莱特(Sewall Wright)与霍尔登(J. B. S. Haldane)把进化建基于健全的统计学理念上,因此,达尔文的学说、基因突变和孟德尔定律之间的伪矛盾被调和。
在20世纪20年代至30年代,出现了一个现代进化综论,把自然选择、基因突变理论、孟德尔遗传定律连结起来,形成普遍适用于生物学任何分支的统一理论。现代进化综论能够通过古生物的过渡化石,以至在发育生物学中的复杂细胞机制,解释在种群的不同物种所观察到的模式。由詹姆斯·沃森和弗朗西斯·克里克在1953年公布的脱氧核糖核酸结构展示了遗传的物理机制。分子生物学提高了我们对基因型和表现型之间的关系的理解。进展也在谱系系统分类学中体现,在对比中绘制过渡 的性状、经发布的可测试框架、和使用进化树。1973年,进化生物学家费奥多西·多布然斯基在一篇文章写下“生物学的一切都没有道理,除非放在进化的光芒之下”,因为它令起初似乎不连贯的自然历史联系起来,转化成连贯的知识体系,形容并预测许多关于在这个星球上的生命所观察到的事实。
此后,现代进化综论得到了进一步的扩展,至解释完整横跨生物层级及综合范围的生物现象(英语:Biological organisation):从基因至物种。此扩展,如进化发育生物学,或简称为进化发生学,强调世代之间的变化(进化)是如何影响生物个体的模式。
遗传
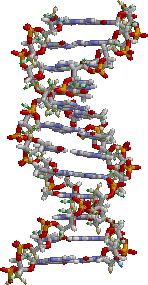
DNA结构,碱基位于中心,外侧环绕着由磷酸根与糖类所形成的双螺旋。
生物体的遗传是发生在一些不连续性状上,也就是生物的特定特征。以人类为例,眼睛的色彩是一个是一项特征,可遗传自父母的其中一个。遗传性状是由基因所控制,而在生物个体基因组中完整的一套基因,则成为基因型。
完整的一套可观察性状,可形成生物的构造或是行为,称为表现型。这些性状来自基因型与环境的交互作用。因此生物体的表现型并非完全来自遗传,例如皮肤的晒黑情况,是决定于个人的基因型与阳光的照射。每个人之所以对阳光有不同的反应,是因为基因型的差异,较显著的例子是拥有白化性状的个体,这类个体不会晒黑,且相当容易晒伤。
基因是DNA分子中一些含有遗传讯息的区域,DNA则是含有四种碱基的长链分子。不同的基因具有不同的碱基序列,这些序列以编码形式形成遗传讯息。细胞里的DNA长链会与蛋白质聚集形成一种生为染色体的构造,染色体上的特定位置,则称作基因座(locus)。有时基因座上的序列在不同个体之间有所差异,这些各式各样变化型态称为等位基因(allele)。突变可使基因序列改变,产生新的等位基因。当突变发生虞基因时,新形成的等位基因可能会影响此基因所控制的性状,使表现型改变。不过单一等位基因对应单一性状的情形较少,多数的性状更为复杂,而且是由许多进行交互作用的基因来控制。
最近的研究结果证实,遗传变化的重要例子不能被DNA的核苷酸序列改变来解释。此类现象被分类为表观遗传系统。DNA甲基化产生染色质,自我持续的代谢循环,通过RNA干扰导致基因表达沉默,蛋白质的三维构象(如朊病毒)的一些区域在部分个体中曾发现表观遗传系统 。
变异
突变
突变是指遗传的物质发生改变。广义的突变包括染色体变异(分为染色体数目和结构变异)和基因突变。不过一般所说的突变,是核酸序列的改变,也就是基因突变。基因突变是产生遗传变异的最根本原因,细胞中的遗传物质(通常是脱氧核糖核酸或核糖核酸)能够经由许多方式改变,例如细胞分裂时的复制错误、放射线的照射、化学物质的影响或是病毒感染。多细胞生物的基因突变,可依照发生的细胞种类分为两种。生殖细胞突变能够遗传到下一代;体细胞突变则通常限制在个体中。
基因突变可能对个体有害,也可能对个体有益,或是两者兼具。有害的隐性基因因为不会出现症状而被保留,当这些隐性基因配成对时,就可能使个体得到病变或是死亡。有一些基因虽然可能会造成病变,但是也可以使个体具有某些优势,例如带有一个镰刀型红血球疾病基因的人,对疟疾更有抵抗力。
对生物个体无益也无害的突变称为中性突变,在种群中的出现频率主要受到突变概率影响。由于这些突变不影响个体的生存机会,因此大多数物种的基因组在没有天择的状况下,依然会有稳定数量的的中性突变不断发生。单一碱基对的变换称为点突变,当一个或多个碱基对插入或是删除时,通常会使基因失去作用。
转座子(transposon)是生物的基因组片段,并且在基因组的进化上扮演重要角色。它们能够移动并插入基因组中,或是取代原有的基因,产生进化上的变异和多样性。DNA复制也被认为每百万年间,会在动物的基因组中产生数十到数百的新基因。
重组
在无性生殖的过程中,染色体上的任何一对等位基因都会一起遗传到下一代。但是对于行有性生殖的物种而言,亲代同源染色体中的等位基因,在制造生殖细胞的减数分裂过程中,会发生基因重组。这是一种不同的脱氧核糖核酸段落断裂并重新组合的过程。
原核生物之间能够透过接合等方式,直接交换彼此的基因,因此重组在原核生物中也比较常见。而较复杂的动物与植物,则通常是在制造生殖细胞的减数分裂时期,因为染色体的互换(crossover)而发生重组。减数分裂重组的发生频率较低,而且排列位置较接近的等位基因,也较不易交换。因此可以由等位基因的重组率计算出基因的相对位置。
此外有性生殖中的孟德尔遗传规则,能够使有害的突变被清除,有益的突变被保留。且因为具有这种清除有害突变效果,因此当一个等位基因无法进行基因重组的时候(例如孤立的Y染色体),便因为有害突变逐渐累积,而使种群的有效种群大小缩减,这种现象称为希尔—罗伯森效应(英语:Hill–Robertson effect))。若是染色体逐渐退化,则称为缪勒氏齿轮(英语:Muller"s ratchet),这种现象比较容易出现在无性生殖的生物中。
进化模型
微观进化与宏观进化
进化可以依据时间长短与差异程度,分成“微观进化”(微进化)与“宏观进化”(广进化)。微观进化指几个世代中,基因频率小范围的变化,例如现今世界各地人类的差异。宏观进化指长时间的进化过程,例如人类与灭绝祖先的关系。宏观进化的历史中可能包括生物群在化石纪录中的突然出现、丢失的环节、物种长期停滞等难以解释的现象。例如寒武纪地层中保存大量化石的现象。
进化速率
关于宏观进化的过程,有许多不同的理论,主要是用来解释化石纪录中难以解释的现象。传统的观念认为,许多微观进化的累积,经历足够时间之后便形成新物种,这样的理论称为渐变论。而渐变论又有许多形式,例如由史蒂芬·古尔德与尼尔斯·艾崔奇在1972年提出的疾变平衡论。这种理论认为生物的进化历史是许多走走停停的过程,在大多数时间并没有太大的变化。而某些个体中存在关键基因(如同源异形盒),可能导致新物种迅速形成并大量繁殖,之后再恢复平衡。如此便能够合理的解释化石纪录不平均的问题。其实古尔德经常以批判渐变论的姿态出现在大众媒体,但是道金斯认为,疾变平衡论只是渐变论的一种形式。而极端的渐变论,认为进化过程是以等速进行,对于化石数量的不平均,则以“化石纪录本身并不完整”来解释。极端的渐变论,仍然经常使用在分子层次,并发展出分子时钟的观念。但是近年的研究发现,分子时钟也并非等速进行,而是在不同物种或是不同年代具有不同的进化速度。且不同的研究中,对分子进化速率的推算差异相当大。
另外有一种跃进论,认为物种只需要一个世代就能够形成,且可能具有创造论与灾变论等形式。创造论与灾变论都是源自宗教,如《创世纪》与大洪水,不过现在的灾变论已将火山爆发、陨石撞击等所有来自地球内外的影响包含在内。
数学模式
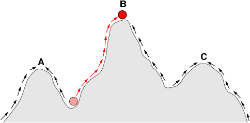
生殖成就空间一般会显现出山峰与山谷,山峰显示生物所能达到的最高生殖成就,山谷则显示其区隔,解释特征差异的存在。生殖成就空间除了可以如此图般以二维方式呈现,也可以三维方式表达。图中直向为生殖成就,横向为表现型变数。
生物的变异可以量化为数字,因此也能够建立出数学模型。较早的数值分类学(numerical taxonomy)将生物的特征量化为数字,并且依照这些数字进行分类,借此找出它们的亲缘关系。
现在的科学家一般认为生物型体的进化,是源自于基因的变异(基因中心进化观点)。而且由于基因突变具有一些规律性,因此复杂的进化过程,可以简化成数学模型。早期的新达尔文主义科学家使用线性的模型,例如将为每一个对偶基因定下一个“天择系数”,借此估计此对偶基因在每一世代中的基因频率。现今则多用非线性的方式分析,例如一种称为生殖成就空间(fitness landscape)的曲面图形,可以用来表达生物个体在繁殖上的能力,与其特征之间的相关性。由图形可以看出具有不同特征的生物可能同时皆有较高的生殖成就。
由于这些生物的变异可以化成许多种不同的变数,因此数学上用来处理多维空间的技巧,便可应用在生物关系的分析。此外这些数学方法的优点是计算过程上相当精确,缺点是其正确性取决于人为的判断与假设。
进化证据
进化的证据可以在系统分类学、生物地理学、比较解剖学、比较胚胎学、古生物学、分子生物学等领域找到。另外进化也可以用来解释抗药性、反祖现象、癌症、马岛长喙天蛾等等。
生物相似性和多元性
生物的多样性和一致性是可以用进化论解释,而智慧创造或直接神创则难以解释这些现象。生物之间的同源特征是进化的证据。因为所有的生物都有共同祖先,所以即使是外观差异非常大的生物,仍然可以在分子生物学、比较胚胎学和比较解剖学上找到相似性。进化也可以解释为什么地球上会有这么多种生物:适应不同栖位。在生物地理学上,相近区域的生物往往在分类学上非常相近,显示出单一物种的辐射进化,而不同地点的生物也可能在类似的环境带有类似的构造,显示出趋同进化。
进化历史的证据
虽然进化有很大部分发生在久远的过去,但是经由痕迹器官、过渡化石、反祖现象和种系发生学重建一个种系或生物器官的进化历史,可以证明过去的生物和现在的生物都经历渐变的进化。
进化论的预测和应用
进化论是可以用来预测或实际应用的,例如细菌的抗药性进化、马岛长喙天蛾的发现都是进化论预测到的;癌症治疗和基于流行病学的公共卫生政策也常用上进化论。这些预测和应用显示进化论是正确的。
机制
遗传变异一方面经由生殖而传递到下一个世代(被称为垂直基因转移),另一方面也可以透过水平基因转移(英语:horizontal gene transfer,简写:HGT),在物种之内或是物种之间传递。尤其是细菌经常使用这种方式交换基因,最近的研究更发现可能有跨物种的水平基因转移存在。基因流(gene flow)则是指基因在生物个体之间转移。
基因型(遗传因子)是产生表现型(外在表现)的根本。而表现型本身也拥有表型可塑性(phenotypic plasticity),能够在基因型未改变的状况下有所变化,并且能够遗传到下一世代。除了基因本身的改变,染色体的重新排列虽然不能改变基因,但是能够产生生殖隔离,并使新物种形成。
一般来说,选择包括了“天择”(自然选择)与“性择”(性选择)。天择的主要原因是物种所居住环境的改变,包括物种之间关系的变化;性择则是物种在繁殖的需求下而产生的选择。而这些性择所留下的性状,可能会有害于个体本身的生存能力。各种选择的分类事实上并不明确,也有一些分类以天择表示所有选择作用,并分为生态选择(ecological selection)与性择。
基因流
基因流也称为迁移(migration),当种群之间并未受到地理或是文化上的阻碍时,基因变异会经由一些个体的迁移,使基因在不同种群间扩散,这样的情形称为基因流。恩斯特·麦尔认为基因流类似一种均质化(homogenising)的过程,因此能够抵销选择适应的作用。当基因流受到某种阻碍,例如染色体的数目或是地理的隔阂,便会产生生殖隔离,这是物种形成的条件之一。
种群中等位基因的自由移动,也受到种群结构的阻碍,例如种群的大小或是地理分布。虽然理想状态中种群的生殖对象完全自由且完全随机,但是现实世界中并非如此,因此地理上的亲近程度会对这些基因的移动造成庞大的影响。而且当迁移数量较少的时候,基因流对进化的影响也较低。
遗传漂变
基因漂变指的是种群中等位基因频率在每一个世代之间的随机的变化。这种变化能够以数学表达,哈蒂-温伯格平衡描述了理想状态情况下(不考虑天择等因素)的数学模型。在理想状态中,后代的等位基因频率将接近随机分布。当种群规模较大,基因漂变的概率会较低;当种群规模较小的时候,基因漂变的现象较为明显。
当一个少数种群从原先种群之中分离而出,且两者的基因频率有所不同,若是分离而出的少数种群与原先种群的基因无法继续交流,则两者的基因频率将渐行渐远。这种现象称为奠基者效应。例如从德国迁移到美国宾夕法尼亚的阿米什人,起源大约仅有200人,且习惯族内通婚。这个种群的埃利伟氏症候群出现频率较其他种群高。
水平基因转移
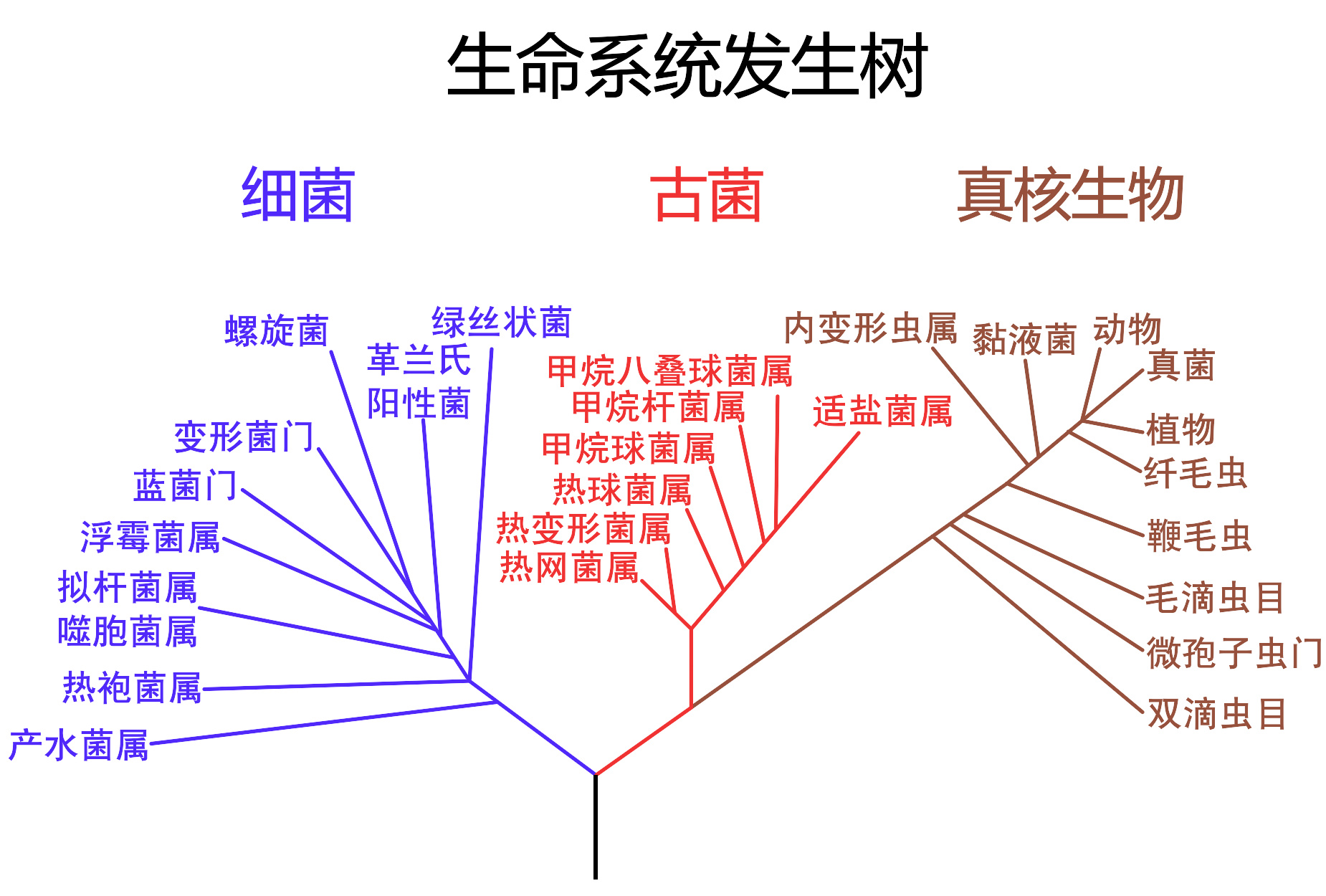
以16SrRNA的基因序列所建立的種系發生樹,將生物演化歷程分為三域系統,包括細菌、古菌與真核生物。第一個提出這種分類的是卡爾·霍斯(Carl Woese)。由於水平基因轉移的存在,使生物的親源關係可能比圖片中所顯示的更為複雜許多
水平基因转移(英语:horizontal gene transfer,简写:HGT),是个体将遗传物质传递到其他非本身后代个体的过程。这种机制使遗传物质得以在无直系关系的个体之间产生基因流。
水平基因转移也可以经由抗原转移(antigenic shift)、基因重整(reassortment)与杂交反应(hybridisation)等现象观察。病毒能够透过转导作用(transduction)在物种间传递基因。细菌则能够与死亡的细菌合体、经由转形作用(transformation),以及与活细菌进行接合(conjugation),而获得新的基因。而新的基因则能够以质体的形式,加入宿主细菌的基因组中。杂交的现象在植物中最显著,此外目前已知还有10种以上的鸟类物种能够杂交。另外在哺乳动物与昆虫中,也有杂交的例子,只是通常杂交后代不具有生殖能力。HGT也是细菌传递抗药性的方式之一,而且有些发现表明HGT是原核生物与真核生物的进化重要机制。
由于HGT的存在,使种系发生学更加复杂,也使早期物种的进化过程出现一种隐藏关系(metaphor)。遗传资讯在生殖作用之外,也能在物种之间传递。这使科学家必须在解释进化关系的时候,表达出物种的隐藏关系,并且将不同的进化历程组合。
自然选择

孔雀显眼又花枝招展的尾羽,是性择的代表性例证,一方面使它容易成为被猎食的目标,另一方面又能够吸引雌性。
由于各种基因的变异,使同一个种群中,不同个体的生存方式和繁殖方式有所不同,当环境发生改变,便会产生天择作用。之所以称为天择,是因为这种选择并非如基因漂变或基因突变一样随机,当环境改变发生时,将只有某些带有特定特征的群体能够通过这些考验。天择有一些特例,有时候被视为与天择拥有相等地位的选择方式。其中包括性择、人择等等。
性择指某个个体因为比起其他个体拥有较高的繁殖机会,因此它们的基因会被保留,使后代继续保有相同的优势。人择指人类为了本身的生存或是喜好而对不同的基因变异进行筛选,通常发生在农业、畜牧业或是宠物的育种上。此外,优生学则是人类对人类所进行的筛选行为。不过人类事实上只是自然界的一部分,因此人择与天择并没有明确的分别。
由于这些特殊的选择机制,导致对于生物适应环境有益的特征,并非在进化过程中一定会出现或是被保留。举例而言,拥有更多的手指对人类的生活可能会更加方便,但是这种方便几乎不会增加任何繁殖机会,甚至反而会减少。
人工选择
狗通过人工选择从狼进化而来。
结果
适应
在天择的作用影响之后,生物能够更加适应它们所处的环境。只要是能够使个体拥有更大生存优势的过程,都可以称为适应。不过需要注意的是,适应并非放诸四海皆准,在一个环境中拥有优势的特征,可能会在另一个环境中成为缺陷,这种现象也能解释为何进化并没有任何预设方向和目的,只有适应或是不适应。
即使“进步”在进化过程中并非必要,但是物种之间的竞争关系,仍能使物种在最基本的环境适应之外,进行更进一步的变化。这些竞争类似人类的军备竞赛,且能够依照物种的关系而分成两类。一种是不对称竞争,指物种受到不同条件的选择,如掠食者与猎物的关系;另一种是对称竞争,指物种受到相同的条件选择,如森林中树木对阳光的争取。此外性择也具有竞争的特性,有些物种在繁殖机会的竞争压力之下,会逐渐发展出相当奇特的外观或行为,例如鲜艳羽毛与吞食异性。
物种形成
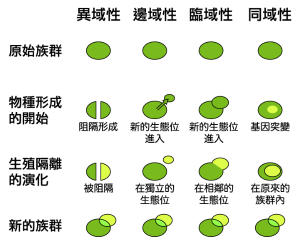
四种物种形成过程的比较图
物种形成受到许多类型的机制影响,主要可以分成2种类型。一种是异域性物种形成(allopatric speciation),发生在种群受到地理隔离,而形成新物种的情况。例如大峡谷两侧的松鼠,由于峡谷地形的阻隔,形成两个物种。不过地理隔离并不一定会使物种分化,当两个物种再度接近,有可能会产生杂交。同域性物种形成(sympatric speciation),是指新物种与固有物种在相同地区生存,却因为繁殖或沟通等行为的差异而产生生殖隔离。例如,印尼一种蝙蝠由于声音不同,而产生三个从未杂交的种群。
另外还有两种较特殊的物种形成方式。近域性物种形成(peripatric speciation)介于异域性与同域性之间,指物种原先受地理隔离,之后因种群扩大而与固有物种接触,却已经形成两个物种。边域性物种形成是由恩斯特·麦尔所提出,这种理论同时支持了疾变平衡理论,且与奠基者效应有关,是小种群作为进化关键的例子。邻域性物种形成(parapatric speciation)指物种虽然生存在相连的区域,但是因为交流的困难而产生新物种。最著名的现象是环状物种(ring species),例如北极圈周围的黑脊鸥(Larus argentatus)。
灭绝

最知名的已灭绝动物之一,渡渡鸟
灭绝指物种或是某个分类上的种群消失,并减少生物多样性。某一物种的最后个体死亡,就是物种灭绝的时刻,即使灭绝前就已经失去了任何繁殖的可能。由于物种的潜在范围可能相当大,因此确定物种灭绝时刻相当困难。
地球上曾经有过多次大规模的灭绝,其原因大多是因为环境,尤其是气候的大幅改变。其中最严重的5次,分别是奥陶纪后期(4亿4千万年前)、泥盆纪后期(3亿6千万年前)、二叠纪后期(2亿5千万年前)、三叠纪后期(2亿1千万年前)与白垩纪末期(6千5百万年前)。其中二叠纪后期的二叠纪灭绝事件,大约95%的海洋生物与70%的陆地动物消失。白垩纪末期的白垩纪灭绝事件,则因为恐龙的灭绝而著名。
生命起源与进化历程

位在美国冰川国家公园的前寒武纪叠层石(stromatolite),可能是已知最早的生命化石纪录。
对于最初始的生命起源,目前尚未明了。而且生物学的进化研究,通常不包括这段初始过程。因为这段过程牵涉到太阳系与地球的形成过程,所以对于生命起源的研究,许多是来自物理学与化学。例如1952年的米勒-尤里实验中,史坦利·米勒与哈罗德·尤里以氨、甲烷、氢气、氰酸与水等分子,模拟地球的原始状态,并首次在实验室中制造出生物的最基本物质。而近年的研究发现,作为氨基酸原料的有机分子,有可能是来自太空中,或是海底火山。
而关于包括遗传物质在内的有机分子进化过程,现在科学家一般认为核糖核酸比蛋白质与脱氧核糖核酸更早出现,之后出现类似反转录酶的蛋白质,最后才有脱氧核糖核酸,不过这些理论的证据并不多。脂肪酸的出现则构成了原始的细胞膜,之后经由内共生等过程,形成最早的单细胞生物。
进化主要事件
在进化过程中,有许多关键性的生物分化,配合地质年代与进化历程,能够归纳出进化时间表。目前已知的化石纪录中,最早生命遗迹是出现在约38亿年前,原核单细胞生物则出现在33亿年前。到了22亿年前,才出现最早的真核单细胞生物,如蓝绿菌。6亿年前藻类与软件无脊椎动物出现。再此之前的年代称为前寒武纪。
古生代是由5亿4千3百万年前到5亿1千万年前所发生的寒武纪大爆发开始,此时大多数现代动物在分类上的门已经出现。之后海中藻类大量出现,而且植物与节肢动物开始登上陆地。最早的维管束植物在4亿3千9百万到4亿9百万年前出现。接着是硬骨鱼类、两栖类与昆虫的出现。3亿6千3百万年前到2亿9千万年前,维管束植物开始发展成大型森林,同时最早的种子植物与爬虫类出现,并由两栖类支配地球。最后爬虫类开始发展,并分化出类似哺乳类的爬虫类,随后发生二叠纪灭绝事件,古生代结束。
中生代开始于2亿4千5百万年前,这时以恐龙为主的爬虫类与裸子植物逐渐支配地球。1亿4千4百万年前到6千5百万年前,开花植物出现,最后中生代结束于白垩纪灭绝事件。
6千5百万年前之后则称为新生代,哺乳类、鸟类与能够为开花植物授粉的昆虫开始发展。开花植物与哺乳动物在这段时间取代了裸子植物与爬虫类,成为支配地球的生物。可能是人类祖先的类人猿出现在360万年前,直到10万年前,现代人(学名:Homo sapiens)才诞生。
现今的研究与应用
进化生物学是研究进化的主要学门,探讨物种的起源和改变,以及物种之间的亲缘关系。这些研究影响了传统的分类学,并导致系统分类学的出现。进化发育生物学(evo-devo)比较不同动物在发育过程中的变化,由此探讨它们之间的关系与进化过程。体质人类学专注于人类的起源与进化,并探讨人种的差异,又称为生物人类学。
为了更深入研究进化过程与机制的细节,许多相关的分支学门产生。例如生态遗传、人类进化、分子进化与种系发生。由于生物学是奠基在其他更基础的自然科学之上,因此数学、统计学、物理学与化学对于了解进化机制也相当重要。例如为基因流、基因漂变等现象提供数学模式的群体遗传学,研究在进化动力影响下,等位基因的分布和改变。
遗传算法则是应用进化与遗传的各种机制,并结合电脑的运算能力来解决许多问题。应用的层面包含工程、设计与通讯科技等。
进化在社会行为研究领域的发展
有些进化研究专注在社会性生物上,称为社会进化学。例如汉弥尔顿提出亲属选择,解释利他行为与的存在。不久之后,艾德华·威尔森出版的《社会生物学:新综合》,解释了社会性生物的各种行为,并在最后讨论套用在人类行为的可能性。
1976年,道金斯在《自私的基因》一书中,认为人类的文化也能以进化解释。他根据基因(gene)这个词,将文化的进化单位称为迷因(meme、在中文也被音译为迷米)。类似作为遗传因子的基因,迷因为文化的遗传因子,也经由复制(模仿)、变异与选择的过程而进化。
社会进化学与迷因学的差异在于,社会进化仍然是一种基因中心观点,以遗传物质分子为天择单位;而迷因学则是以非基因的文化为天择单位。
误解
进步、复杂化与退化
有些物种(如人类),常被认为是比其他的物种更高级,甚至是进化的方向与目的所在。且认为进化的过程必定会使生物愈来愈复杂,或是进行与进化相反的退化。而现在的生物学家认为进化是没有方向的过程,也没有任何预先计划的目标。虽然在已知的进化过程中,确实具有逐渐复杂的现象,但是依然有许多物种保持在较简单的状态,如细菌。因此复杂性可能增加也可能减少,或是维持不变,结果取决于天择的机制。
物种形成
物种形成有时候被认为是无法直接观察的现象,并得出进化是不科学的结论。但是科学的发现不仅是经由可重复的实验,均变说(uniformitarianism)使科学家得以用经验来推论事物的原因。此外物种形成的例子也出现在植物。还有刺鱼(stickleback)的外胚叶发育不全(ectodysplasin)等位基因,被用来当作研究基因转变与物种形成的模型。有一种类似的观点,认为微观进化是可以观察,而宏观进化则无法观察。但是由于宏观进化的机制与微观进化相同,所以宏观进化事实上已经在微观进化中被观察。而且物种之间基因序列的比较,也显示少量的遗传变异,就可以导致外表相当大的变化。
熵与生命
有些观点认为进化增加复杂性的情形,违反了热力学第二定律。熵是物理学上的参量,这个定律是指在一个孤立系统中,熵只会增加或是维持不变,可以使用的自由能逐渐减少,最终反应逐渐趋于动态平衡。这种观点忽略了生态系事实上并非闭合系统,所有生态系中生物所获得的自由能,都是来自太空中,尤其是太阳。太阳、地球与太空的系统并不违反热力学第二定律,因为太阳与地球辐射所产生的自由能,远超过生物进化所需。

讽刺漫画反映了早期人们对于“人类与猿类具有共同祖先”这个观念的抵触。
政治
许多政治或宗教领袖以为进化论在学界无共识、创造论和进化论一样有效。如前总统里根曾在1980年的一场竞选活动中表示:“进化是一个理论,只是一个科学理论,直到现在依然在科学界中受到挑战,并且尚未被科学界认为绝对正确。”这类说法强调进化只是一个理论,所以并不是真实存在的事物。然而对科学家而言,理论并非与事实对立。事实是指经验上所得的资料或数据,理论则是对事实的解释与想法。里根更说,如果学校要教进化论,也要教圣经上对人类来源的解释,“而近年来的发现,指出了进化论重大的破绽”。这正与事实相反。2005年美国总统布希公开赞成学校同时教授智慧设计论与进化论。批评者说:“乔治·布什可以证明,受过哈佛和耶鲁教育,也影响不了一个人的顽固。”。
2005年的美国宾州多佛学区案中,法官判决学校在2004年开始教授的智慧设计论违反法律,其它学区因为避免官司而从此放弃教授此论。。2010年,美国宗教学院定下指导方针,智慧设计论不应在科学课中教授,只可作为文学或社会科学的课程。
宗教
自从《物种源起》出版之后,进化论在宗教的争议就持续不断。最大的争议,便是关于人类进化的部分,与《圣经》中《创世纪》的冲突。
进化论的出现,也导致一些以宗教观点解读进化论的理论出现。例如天主教,将其信仰与进化论调和为神导进化论。而除了传统的创造论之外,也有一种称为智慧设计的理论出现,是认为生物的出现必定受到某种智慧体的安排。
伦理
另一种对于进化论的批评,对象则是社会达尔文主义,以及衍生而出的种族主义、优生学与生育控制等19世纪末与20世纪初的产物。这类思想主要是起源於哲学家与政治人物对达尔文主义的借用,例如最早提出类似理论的赫伯特·史宾赛。而进化学者中也有一些本身具有优生学与种族主义等思想,例如提出胚胎重演论的海克尔。
参看
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

相关资料





- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读



关于我们

APP下载



{{item.time}} {{item.replyListShow ? '收起' : '展开'}}评论 {{curReplyId == item.id ? '取消回复' : '回复'}}