





-
 QQ空间
QQ空间
-
 QQ好友
QQ好友
-
 微信好友
微信好友
-
 新浪微博
新浪微博
恐爪龙属
描述
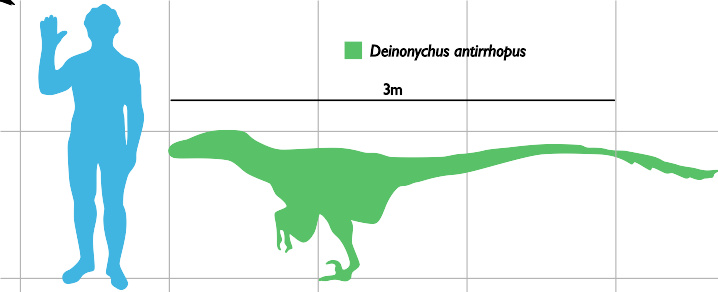
人类与恐爪龙的比例。
根据恐爪龙的最大型标本,恐爪龙的体型可达3.4米,头颅骨最大可达41公分长,臀部高度为0.87米,而体重最高可达73公斤。它的头颅骨有强壮的颌部,有约60根弯曲、刀刃形的牙齿。奥斯特伦姆最初将恐爪龙的不完全颅骨,重建成三角形、宽广的头部,类似异特龙。在发现更多、更完整的恐爪龙与其近亲的化石后,显示恐爪龙的上腭部较呈拱形,口鼻部较狭窄,颧骨宽广,使头部看起来较为立体,与奥斯特伦姆的重建不同。恐爪龙颅骨与伶盗龙不同,头顶较坚固,类似驰龙;伶盗龙的鼻部上侧往下凹,这点与恐爪龙不同。头颅骨及下颌都有洞孔,可减轻头部重量,而恐爪龙的眶前孔是特别的大。按头颅骨来推算,眼睛主要是向两侧的。
就像其他的驰龙科,恐爪龙有大型手掌与三根手指。第一指最短,而第二指最长。每只后肢的第二趾都有镰刀状的趾爪,长度约12公分,有可能是作为捕猎动物。它可以先向前戳刺,并向下割来撕破猎物。相对于恐爪龙的体型,这些趾爪来的相当地大。恐爪龙的身体是靠尾椎及人字骨,在高速转向时来维持稳定及平衡。
分类
恐爪龙是最著名的驰龙科恐龙之一,且是伶盗龙的近亲,伶盗龙的体型较小,生存于上白垩纪的蒙古。它们的都属于伶盗龙亚科,这个亚科最初是由瑞钦·巴思钵(Rinchen Barsbold)于1983年所提出,而当时只有单一的伶盗龙属。后来菲力·柯尔(Phil Currie)将大部分的驰龙科都放在这亚科中。生存于上白垩纪蒙古的白魔龙,以及同时代在北美洲的蜥鸟盗龙都可能是恐爪龙的近亲,但后者因化石较少而难以分类。伶盗龙及其近亲都是使用第二趾爪作为猎杀工具,而不像驰龙是以厚实的头颅骨来作为武器。驰龙科与伤齿龙科共同组成恐爪龙下目,是鸟类的姊妹分类单元。在种系发生学上,恐爪龙下目代表了一群不属于鸟类,却与鸟类是近亲的恐龙。
发现与命名

平衡恐爪龙的骨架模型

编号 YPM 5205 的正模标本(左后肢)重建模型
恐爪龙的化石发现于美国蒙大拿州与怀俄明州的Cloverly组,以及奥克拉荷马州的鹿角组。Cloverly组的地质年代为白垩纪早期的阿普第阶晚期至阿尔布阶早期,距今约1亿1500万到1亿800万年前。此外,马里兰州大西洋沿岸平原地带的波多马克组,发现了一些可能属于恐爪龙的牙齿,年代为阿普第阶。
恐爪龙的第一副化石,是在1931年由巴纳姆·布郎(Barnum Brown)所带领的队伍在蒙大拿州南部的Cloverly组发现。布郎当时主要是想发掘并处理腱龙的遗骸,但在他交予美国自然史博物馆的1931年报告中,指出发现了一小型的肉食性恐龙,发现位置接近腱龙化石,但因陷在石灰岩而难以作清洁处理。他非正式地将之命名为"Daptosaurus",并准备描述及展示其骨骼,但他却没有完成。另外,布朗将一个恐爪龙的牙齿,与另一种较小型兽脚类恐龙的骨骸混淆在一起,并非正式命名为"Megadontosaurus"。
约30年后,从1964年8月开始,古生物学家约翰·奥斯特伦姆(John Ostrom)率领一个耶鲁大学皮博迪自然史博物馆的挖掘团队,在其后的两年内发现了超过1000个骨头,来自于至少三个个体。因为很难判断这些化石的正确位置,所以恐爪龙的正模标本(编号YPM 5205)只限于完整的左足部,与部分的右足部,但都确定属于同一个体。其他标本则在耶鲁大学的皮博迪自然史博物馆区分、标示成50个不同的项目。
在接下来几年,奥斯特伦姆及格兰特·迈耶(Grant E. Meyer)研究这些新发现化石,以及布朗所命名的"Daptosaurus"化石,发现它们是同种生物,并在1969年将这些化石命名为平衡恐爪龙(Deinonychus antirrhopus)。种名antirrhopus意为“平衡”,指的是它们坚挺尾巴的可能用途。同年,奥斯特伦姆公布更广泛的恐爪龙专题论文。
奥斯特伦姆同时重新检验布朗所命名的"Megadontosauru",他发现那只牙齿是属于恐爪龙,而骨骼则是来自另一类动物。他在1970年,将骨骼部分命名为小猎龙。
虽然在1969年发现大量的恐爪龙化石,但许多重要部位的骨头并没有发现,剩下的难以确认。这些化石包含少数眼眶后颅骨,没有股骨、没有荐骨、没有叉骨或胸骨、也没有脊椎,以及一个被奥斯特伦姆认为是鸟喙骨的小型碎片。奥斯特伦姆将恐爪龙重建成具有独特的骨盆,耻骨呈梯形且平坦,与坐骨长度一样。
更多发现

平衡恐爪龙的现代想像图,羽毛是参考相关物种而来的。
在1974年,奥斯特伦姆公布了另一份恐爪龙专题论文,并指出他原本鉴定的耻骨,其实是个肩膀上的喙骨。同一年,法里什·詹金斯(Farish Jenkins)所率领的哈佛大学挖掘团队,在蒙大拿州发现了另一个恐爪龙标本。这个新标本增添了七个没发现过的部位,包含:股骨、耻骨、荐骨、肠骨、以及脚掌与跖骨。奥斯特伦姆研究了这个新标本,并推翻他之前的恐爪龙重建。这次恐爪龙被重建成具有长的耻骨,而奥斯特伦姆并推论耻骨稍微后倾,类似鸟类。
在纽约的美国自然史博物馆,有一个展览中的恐爪龙骨架,上有最初的标本组成。哈佛大学的比较动物博物馆则有其他的标本展览中。它们的恐爪龙骨架都缺乏胸骨、胸肋、叉骨、腹肋。耶鲁大学也有恐爪龙骨架,但与前两者的发现处不同。
在奥斯特伦姆研究了的正模标本后,仍有几个陷在石灰岩的小型标本未被处理,而摆放在美国自然史博物馆。这些标本大部分是独立的骨头及骨头碎片,包括了标本埋葬时的周围母岩。在2000年时,Gerald Grellet-Tinner与彼得·马克维奇(Peter Makovicky)重新研究这些标本,发现一些有趣及被忽略的特征。当中的几条修长骨头,原先被认为是骨化肌腱,可使恐爪龙尾巴坚硬,其实是腹肋。而且在原先恐爪龙标本的周围,也发现了一些被忽略的蛋壳化石
在后来的详细报告中,Tinner与马克维奇提出这些蛋几乎肯定是属于恐爪龙的,是首批发现的驰龙科蛋。再者,其中一个蛋壳的外表面紧接恐爪龙的腹肋,可见恐爪龙可能正在孵蛋。由此推断恐爪龙是用身体的热量来孵蛋,类似现今鸟类,这也显示它们是温血动物。在2007年,格里高利·艾利克森(Gregory Erickson)与其同事的进一步研究,发现正模标本的年龄约13或14岁,而且是稳定的成长。同一份研究中也指出,恐爪龙在死亡前便已停止成长,与其他已知的兽脚类恐龙不同。
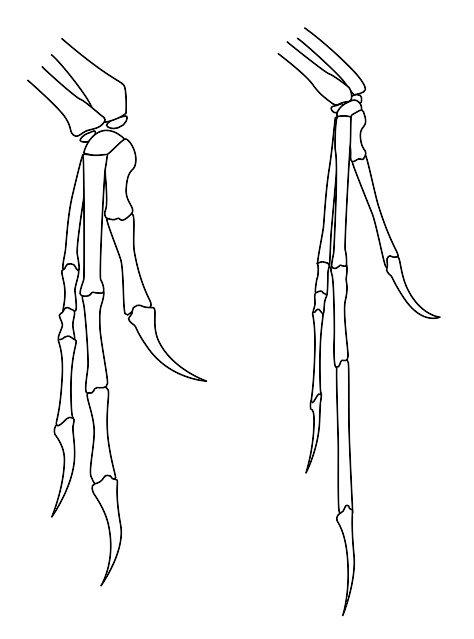
恐爪龙的手(左)与始祖鸟的手(右)比较。
影响
奥斯特伦姆在1969年的恐爪龙研究,被认为是20世纪中期最重要的恐龙发现。因为这个新发现的活跃、敏捷恐龙,改变了很多关于恐龙的科学研究,以及一般大众对恐龙的认知,且开启了对恐龙可能是温血动物的辩论。这个认知的变化称为恐龙文艺复兴。几年后,奥斯特伦姆发现了恐爪龙与鸟类的手掌的相似性,使他认为鸟类是从恐龙所演化而来。三十年后,这个观念几乎是被普遍地接受。
目前还没有发现恐爪龙具有羽毛的证据。但某些驰龙科化石已发现有羽毛的直接与间接证据,显示这个演化支普遍具有羽毛。生存年代较早、较为原始的小盗龙,也属于驰龙科,已有多个标本被发现具有羽毛痕迹,分布于前肢、腿部、尾巴,羽毛形态已经相当类似现代鸟类。另一种在中国发现的中国鸟龙,化石也被发现有类似羽毛的结构,其特征显示它们较恐爪龙更为原始,其生存年份也早于恐爪龙。在2007年,恐爪龙的近亲伶盗龙被发现前肢尺骨具有羽茎瘤。现代的鸟类也具有羽茎瘤,是羽毛的附着处。这个发现显示伶盗龙也具有羽毛。在其他兽脚亚目中,如尾羽龙、鸟面龙、北票龙及帝龙,也有发现羽毛的轮廓,根据亲缘分支分类法的位置,更令人相信所有驰龙科都是有羽毛的。菲力·森特(Phil Senter)在进行恐爪龙、斑比盗龙的前肢活动范围研究时,提出驰龙类可能都具有飞羽。
古生物学
栖息地
地质学证据显示恐爪龙栖息于泛滥平原或沼泽。Cloverly组及鹿角组的古生态环境包含有热带与副热带森林、三角洲及潟湖,并类似今日的路易斯安那州。
其他同时期的恐龙包括有草食性的楯甲龙、鸟脚下目的西风龙及腱龙。在奥克拉荷马州,恐爪龙的生态系统也包括了巨大兽脚亚目的高棘龙、蜥脚下目的波塞东龙、棱角鳞鳄、以及雀鳝目的Lepisosteus。如果马里兰州所发现的牙齿是属于恐爪龙的,那恐爪龙也在当地区与星牙龙、伤龙、孔牙龙(目前只发现牙齿)等恐龙共同生存。Cloverly组中段的地质年代约为1亿1500万到1亿800万年前。
猎食行为

集体攻击的恐爪龙
在Cloverly组,常在腱龙化石的附近发现恐爪龙的牙齿。而在其中两个采石场,已发现相当完整的恐爪龙化石,就位在腱龙化石的附近。第一个是蒙大拿州Cloverly组的耶鲁采石场,发现过四个恐爪龙的成年个体与一个幼年个体,以及众多的牙齿。基于在同一位点发现大量恐爪龙的骨骼,且在腱龙的附近发现恐爪龙的牙齿,估计恐爪龙是猎食腱龙的。奥斯特伦姆与德斯蒙德·麦斯威尔(W. Desmond Maxwell)更以此推断恐爪龙是成群生活及猎食的。第二个采石场则位在奥克拉荷马州的鹿角组。该地发现六个腱龙的部分骨骸,体型不一,附近另有一个恐龙的部分骨骸与众多牙齿。一个腱龙的肱骨上有齿痕,可能是恐爪龙所留下。在1998年,丹尼尔·布林曼(Daniel L. Brinkman)等人提出成年的恐爪龙为70到100公斤,而成年腱龙为1到4公吨。单一的恐爪龙无法猎杀成年的腱龙,显示它们应是采群体方式猎食腱龙。
但是在2007年,布林曼与B. T. Roach基于现今肉食性动物的猎食方式及腱龙位点的埋葬学,质疑了恐爪龙在群猎时的合作。现今的主龙类(鸟类及鳄鱼)及科莫多龙有少数的合作性捕猎,不过它们主要是单独猎食,或是被吸引至已死的尸骸,而会为了抢食猎物尸体而发生打斗。例如,当一群科莫多龙一同摄食时,最大的会先吃,且会攻击其他尝试吃食的较小型科莫多龙;如果小型的科莫多龙被杀死,它们的尸体会被同类抢食。当这些资料套用在腱龙与恐爪龙的位点时,这似乎与科莫多龙与鳄鱼的摄食模式很吻合。在这些位点发现的恐爪龙骨骼都是接近成年的,而失去了的部位可能是被其他恐爪龙吃了。在2007年,科学家们在中国山东省发现了第一个大量的驰龙科足迹化石。这个足迹化石可能是由大型驰龙科恐龙所留下,而且是由六个相近大小的个体,沿者海岸共同前进。这些个体相距约一米,朝同一方向缓慢前进。研究人员提出,虽然该足迹化石并不能明确指出驰龙科的猎食行为,但无法排除它们以群体猎食的可能性。

恐爪龙用脚掌制伏一只西风龙

恐爪龙的头颅骨
在2011年的一项恐爪龙下目趾爪研究,比对不同恐爪龙类、现代鸟类的脚掌结构、趾爪形状,根据这些现代鸟类的猎食行为,分析这群恐龙的压制猎物能力。这项研究认为伶盗龙等驰龙类可能以小型猎物为食。研究发现,许多掠食性现代鸟类也有大型第二趾爪,当它们猎食小型猎物时,会使用脚掌、趾爪将猎物固定在地面上,用体重施压于猎物,再用鸟喙吞食这些猎物,尤其是鹰科。这项研究推论驰龙类会以类似的方式捕抓、固定猎物,并开始吞食猎物的身体,直到猎物失血过多、失去生理机能。研究发现驰龙类的后肢、脚掌结构最类似鹰,也有大型第二趾爪、类似的脚掌抓取范围;驰龙类的跗跖骨短、脚掌力量,则比较类似猫头鹰。伶盗龙的颌部、前肢结构,也符合这项研究的结论。驰龙科的前肢强壮,但极可能覆盖者羽毛,它们吞食猎物时可能会将前肢张开,以伺机保持身体的平衡;尾巴也具有平衡身体的功能。驰龙科的颌部结构虚弱,可能主要用来吞食、撕咬猎物的肉体,而非以快速、强壮力量攻击猎物。研究人员推测,近鸟类可能共同拥有这些演化特征,而现代鸟类则继承这些特征。
咬合力
在2005年,一群科学家重建恐爪龙的颌部肌肉群,计算恐爪龙的咬合力,发现它们的咬合力相当于现代美洲鳄的约15%。在2010年,另一群科学家研究一个腱龙化石上的恐爪龙齿痕,发现这个恐爪龙是个大型个体,而且能直接咬穿大型动物的骨头。他们计算出的恐爪龙咬合力,高于之前的生物力学研究结果,介于4,100到8,200牛顿,这个数值大于许多现代肉食性哺乳动物,接近类似体型的鳄鱼。
这群科学家也发现,恐爪龙的齿痕非常少见,而大型兽脚类恐龙的齿痕则较为常见,例如暴龙。他们推论,恐爪龙可能很少在进食时接咬穿骨头,恐爪龙可能在猎捕猎物时使用高咬合力、或是在抵抗更大型的掠食动物时。
四肢功能

恐爪龙的脚掌与镰刀趾爪
恐爪龙的最著名特征是镰刀状的第二趾爪,但不同标本的第二趾爪的形状与弯曲度都有所不同。奥斯特伦姆在1969年研究的正模标本,其第二趾爪是非常弯曲的,而在1976年研究的标本则较直,且与其他的趾爪很相似。奥斯特伦姆认为这只镰刀状趾爪的大小及形状,随者个体、性别或年纪的不同,而有所不同。目前已发现驰龙科在行走时,第二脚趾后缩、离地的解剖学与足迹化石证据。
奥斯特伦姆猜测恐爪龙是以前肢捉住猎物,并以镰刀状趾爪来割破肚子。某些人员更提出,恐爪龙的镰刀状趾爪可划开大型角龙类的身体。但是后来的研究提出,这些镰刀爪并非用作割开猎物的,而是刺戳猎物的。另有研究人员参考恐爪龙、伶盗龙的脚部结构,使用机械模型攻击猪尸体,但只能在尸体刺出浅穴,无法有效划开肉体。研究人员假设,镰刀状趾爪也可能用来爬上动物的身体。
在1970年,约翰·奥斯特伦姆曾参考鸵鸟与食火鸡,发现这些鸟类会用较大型的第二趾爪攻击猎物,进而产生恐爪龙会利用第二趾爪攻击猎物的理论。食火鸡的第二趾爪可长达12.5公分。奥斯特伦姆并引用一个1958年的鸟类论文,认为后肢与第二趾爪,可用来将猎物的身体撕开、将内脏挖出。在近年,Chhristopher P. Kofron研究241件食火鸡的攻击纪录,发现总计有1个人类、2只狗的被攻击死亡案例,但没有证据显示食火鸡会用第二趾爪将猎物的内脏挖出。相反地,食火鸡将趾爪作为防卫武器,攻击对它们产生威胁的动物、或威吓对方叫鹤也具有较大的第二趾爪,却是用在撕裂小型猎物,以适合吞咽。。在2011年的一项恐爪龙类趾爪研究,也认为第二趾爪无法作出有效的刺穿、划开猎物身体的动作,并提出第二趾爪有固定猎物功能的理论。
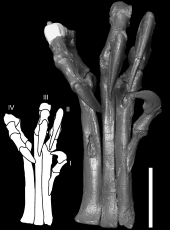
收缩状态的脚掌(编号MOR 747)
在2002年,肯尼思·卡彭特(Kenneth Carpenter)的生物力学研究,确认了恐爪龙的前肢最可能是用来捉住猎物的,因为其前肢的长度较其他兽脚亚目更易于捕捉。大及长的鸟喙骨显示,恐爪龙有强壮的前肢肌肉,更加强了这个解释。卡彭特的研究也制作骨头模型来实验。实验证明恐爪龙是不能掘曲它的手臂,像鸟类般紧贴身体,这与嘉克斯·高斯特(Jacques Gauthier)及葛瑞格利·保罗(Gregory S. Paul)两人在80年代的各别推论有所不同。
在2006年,菲力·森特(Phil Senter)指出恐爪龙的前肢未必只用来捕捉猎物,也可以将物体抓向前方。恐爪龙被认为极可能前肢具有羽毛,但这会妨碍前肢的活动范围。例如,当恐爪龙将前肢往前伸直时,掌心会自动翻转朝前。当两个前肢同时伸直时,前肢的羽毛会妨碍行动。所以森特认为,当恐爪龙在同时间只能用一只手抓近物体,而不能同时用双手。羽毛可能也限制了手指的功能,所以可能只有第三指可发生作用,例如在探挖土中的小型猎物时,也只有第三指可做出90度的弯曲。在2001年,Alan Gishlick也对恐爪龙的前肢力学做了研究,他认为即使前肢具有羽毛,它们并不会明显影响前肢的抓取能力。而且,当手掌抓取时,可与前肢作90度垂直,并可做类似拥抱的动作。这些发现与卡彭特与森特的前肢研究一致。
在2006年,克里斯登·帕森斯(Kristen Parsons)提出恐爪龙的幼年及接近成年标本,与成年的标本有着一些形态上的不同。例如,幼年个体的手臂在比例上较成年的为长,显示可能幼龙与成年的行为模式可能也不同。另一个证据是脚趾趾爪的功能。帕森斯提出幼年个体的趾爪比较弯曲,可能有助于翻爬树干;成年个体的趾爪比较笔直,有助于在地面移动。这个理论是根植于小型驰龙科可能会使用趾爪在攀爬树干。
速度
驰龙科,尤其是恐爪龙,一直都被大众媒体认为是奔走快速的恐龙,而奥斯特伦姆亦在原先的描述中有此推论。但是在1969年首次描述时,并没有发现完整的恐爪龙后脚,而奥斯特伦姆所估计的股骨长度,后来被证实是过长。奥斯特伦姆在1976年的研究注意到,在评估速度上,脚掌与胫骨的相对长度,比股骨与胫骨的比例更为重要。现今奔走快速的鸟类,如鸵鸟,脚掌与胫骨比例为0.95;而在恐龙中,似鸵龙是0.68,恐爪龙则是出奇的低,只有0.48。他于是结论出恐爪龙相对于其他恐龙并不怎么快速,且更不是现今不能飞的鸟类的对手。
恐爪龙的脚掌/胫骨比例低,部分是因它非常短的跖骨。小型个体的脚掌/胫骨比例,仍比较大个体的为长。奥斯特伦姆认为短的跖骨是与镰刀爪的功能有关,当年纪渐长时跖骨的比例会更为短小,更支持他的理论。他解释所有这些特征(短的二趾上有大的爪、短的跖骨等)都是为使后肢成为攻击性的武器,镰刀爪可以向上及向下攻击,而脚则同时向后及向下拉,把猎物割破及撕开。奥斯特伦姆指短的跖骨减少了脚骨在攻击时的整体压力,而恐爪龙的特殊肌肉构造是用作猎食而非奔跑或行走。所以他断定恐爪龙的脚是平衡奔跑,及减低作为武器时遭受的压力。
在1981年,Richard Kool对于加拿大恐龙足迹化石的研究中,他针对英属哥伦比亚盖森组所发现的不同物种的足迹,做了初略的行走速度估计值。其中一个可能是恐爪龙留下的足迹,被估计以时速10.1公里行走。
蛋化石

躺卧的平衡恐爪龙
在2000年,从正模标本周遭石块鉴定出来的恐爪龙蛋,可让研究人员与其他兽脚亚目的蛋结构、巢穴及繁殖作出比较。在2006年,Tinner与马克维奇评估这些蛋壳标本的时候,他们提出不同的假设,例如驰龙科是吃蛋的,或者蛋碎片与该只恐爪龙是碰巧在一起的。因为这些蛋壳是在恐爪龙的腹肋与前肢之间,并非是在胃部之内,故这些蛋并非它们的食物。而根据蛋壳的挤压与破碎状况,可推论蛋是完整埋入土中的,而在化石化的过程中破裂。另外,由于蛋周围的骨骼并非分散或分开的,反而是与在生时差不多的姿势,可见这个位点在化石化期间并没有受到影响,故巧合之说亦不成立。事实上,腹肋通常很难发现保持天然状态的,可见这些蛋是在埋葬时就在恐爪龙的腹下。这可能是恐爪龙的孵蛋或筑巢行为,像伤齿龙科或窃蛋龙科;或是该恐爪龙死时,蛋仍在输卵管之中。
根据这些蛋的微结构,发现它们是属于兽脚亚目,因为它们有着其他兽脚亚目蛋的特征,并与鸟臀目及蜥脚下目蛋的不同。与其他手盗龙类比较,这些蛋化石较像窃蛋龙科的蛋,而非伤齿龙科的蛋。虽然该蛋经已碎裂,而很难评估其大小,但Tinner与马克维奇根据蛋所经过的骨盆腔,推测出蛋的直径约为7厘米。这个大小与最大的葬火龙蛋的直径7.2厘米接近。而葬火龙与恐爪龙的体型接近,支持了蛋的估计值。再者,葬火龙蛋壳及恐爪龙蛋壳厚度差不多完全一致,由于蛋壳厚度影响着蛋的体积,故此有指它们蛋的大小是完全一样的。
病理
在2001年,布鲁斯·罗斯柴尔德(Bruce Rothschild)等人发表一份兽脚类恐龙的压力性骨折研究。他们研究恐爪龙的43个手掌骨头、52个脚掌骨头,没有发现压力性骨折的迹象。
编号YPM 5205标本的第二脚趾第二趾骨,被发现有骨折后愈合的迹象。
大众文化
自从恐爪龙被发现并描述之后,就一直频繁地作为驰龙科的代表物种出现在各种科普读物和电视电影当中。恐爪龙出现在探索频道的纪录片《恐龙战争》中,片中两只恐爪龙齐心协力杀死了亚成年的波塞东龙。现实中,恐爪龙与波塞东龙并没有证据表明它们之间存在捕食关系。在2008年的纪录片《侏罗纪格斗俱乐部》中,一群恐爪龙杀死一只肌腱龙。
至于电影《侏罗纪公园》系列中伶盗龙(Velociraptor)却类似恐爪龙的表现,后来在续集《侏罗纪世界》剧中借负责恐龙基因的吴博士之口解释,复制恐龙是迎合人们对恐龙印象制作出来的,并非原本样貌。
参考资料
^Ostrom, J. H. (1970) "Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana", Bulletin of the Peabody Museum of Natural History 35:1–234.
^(英文)Norell, Mark A.; & Makovicky, Peter J.Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis. American Museum Novitates. 1999, 3282: 1–45.
^3.03.1Ostrom, J. H. Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana. Bulletin of the Peabody Museum of Natural History. 1970, 35: 1–234.
^4.04.14.2(英文)Paul, G.S. Predatory dinosaurs of the world : a complete illustrated guide. New York: Simon and Schuster. 1988: 366–369. ISBN 0-671-61946-2.
^5.05.1(英文)Maxwell, W. D. & Witmer, L. M. New Material of Deinonychus (Dinosauria, Theropoda). Journal of Vertebrate Paleontology. 1996, 16 (3): 51A.
^(英文)Witmer, Lawrence M. & Maxwell, William D. The skull of Deinonychus (Dinosauria:Theropoda): New insights and implications. Journal of Vertebrate Paleontology. 1996, 16 (3): 73A.
^7.07.1(英文)Norell MA, Makovicky PJ. Dromaeosauridae. (编) Weishampel DB, Osmólska H, Dodson P. The Dinosauria (2nd Edition). University of California Press. 2004: 196–210. ISBN 978-0-520-24209-8.
^8.08.18.2(英文)Norell, M.A.; Clark, J.M., Turner, A.H., Makovicky, P.J., Barsbold, R., and Rowe, T. A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia). American Museum Novitates. 2006, 3545: 1–51.
^(英文)Turner, A.S.; Hwang, S.H.; and Norell, M.A.A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia(PDF). American Museum Novitates. 2007, 3557: 1–27.
^(英文)Barsbold, R. Carnivorous Dinosaurs from the Cretaceous of Mongolia. The Joint Soviet-Mongolian Palaeontological Expedition, Transactions. 1983, 19: 1–117.
^(英文)Currie, P. J. New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda). Journal of Vertebrate Paleontology. 1995, 15 (3): 576–591. doi:10.1080/02724634.1995.10011250.
^Paul, G.S. Predatory dinosaurs of the world : a complete illustrated guide. New York: Simon and Schuster. 1988: 358. ISBN 0-671-61946-2.
^(英文)Benton, Michael J.Vertebrate Palaeontology (Third Edition). Blackwell Publishing. 2004: 472. ISBN 9780632056378.
^(英文)Ostrom, J. H. Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana. Bulletin of the Peabody Museum of Natural History. 1970, 35: 1–234.
^(英文)Brinkman, D. L.; R. L. Cifelli, and N. J. Czaplewski. First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian-Albian) of Oklahoma. Oklahoma Geological Survey Bulletin. 1998, 146: 1–27.
^16.016.1(英文)Chen, Z.-Q.; Lubin, S. A fission track study of the terrigenous sedimentary sequences of the Morrison and Cloverly Formations in northeastern Bighorn Basin, Wyoming. The Mountain Geologist. 1997, 34: 51–62.
^17.017.1(英文)Burton, D.; Greenhalgh, B.W.; Britt, B.B.; Kowallis, B.J.; Elliott, W.S.; Barrick, R. New radiometric ages from the Cedar Mountain Formation, Utah and the Cloverly Formation, Wyoming: implications for contained dinosaur faunas. Geological Society of America Abstracts with Programs. 2006, 38 (7): 52.
^(英文)Lipka, T. R. The Affinities of the Enigmatic Theropods of the Arundel Clay Facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland. (编) Lucas, S.G., Kirkland, J.I. and Estep, J.W. Lower and Middle Cretaceous Terrestrial Ecosystems. New Mexico Museum of Natural History and Science Bulletin, 14. Albuquerque: New Mexico Museum of Natural History and Science. 1998: 229–234. OCLC 40283894.
^19.019.119.219.3(英文)Grellet-Tinner, G.; and Makovicky, P. A possible egg of the dromaeosaur Deinonychus antirrhopus: phylogenetic and biological implications. Canadian Journal of Earth Sciences. 2006, 43: 705–719.
^20.020.1(英文)Norell, M. A.; Gaffney, E. S.; and Dingus, L. Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf. 1995: 126–130. ISBN 0-679-43386-4.
^21.021.121.221.321.4(英文)Ostrom, J. H. Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Peabody Museum of Natural History Bulletin. 1969, 30: 1–165.
^Ostrom, J.H., 1969, "A new theropod dinosaur from the Lower Cretaceous of Montana", Postilla128: 1-17
^(英文)Ostrom, John H. The Pectoral Girdle and Forelimb Function of Deinonychus (Reptilia: Saurischia) : A Correction. Postilla, Peabody Museum of Natural History Bulletin. 1974, 165: 1–11.
^(英文)Ostrom, J.H.On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus. Breviora. 1976, 439: 1–21.
^(英文)American Museum of Natural History.Deinonychus.. American Museum of Natural History. 2007 [2007-07-13].
^(英文)Makovicky, P. J.; Grellet-Tinner, G. Association between a specimen of Deinonychus antirrhopus and theropod eggshell. In Bravo, A.M. and T. Reyes (eds.) t international symposium on dinosaur eggs and babies,Isona i Conca Dellà Catalonia, Spain, 23–26 September 1999. 2000: 123–128.
^(英文)Grellet-Tinner, Gerard. Oology And The Evolution Of Thermophysiology In Saurischian Dinosaurs: Homeotherm And Endotherm Deinonychosaurians?. Papeis Avulsos de Zoologia. 2006, 46 (1): 1–10. doi:10.1590/S0031-10492006000100001.
^(英文)Erickson, Gregory M.; Curry Rogers, Kristina; Varricchio, David J.; Norell, Mark A.; and Xu, Xing.Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition(pdf). Biology Letters. 2007, 3 (5): 558–61 [2007-07-26]. PMC 2396186. PMID 17638674. doi:10.1098/rsbl.2007.0254.
^(英文)Fastovsky DE, Weishampel DB. Theropoda I:Nature Red in Tooth and Claw. (编) Fastovsky DE, Weishampel DB. The Evolution and Extinction of the Dinosaurs (2nd Edition). Cambridge University Press. 2005: 265–299. ISBN 978-0-521-81172-9.
^(英文)Ostrom, J. H. Archaeopteryx and the origin of birds. Biological Journal of the Linnean Society. 1976, 8: 91–182.
^31.031.1(英文)Xu, X.; Zhou, Z.; Wang, X.; Kuang, X.; Zhang, F. & Du, X. Four-winged dinosaurs from China. Nature. 2003, 421 (6921): 335–340. PMID 12540892. doi:10.1038/nature01342.
^(英文)Hwang, S. H.; Norell, M. A.; Ji, Q. & Gao, K.al.2002.pdf New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China(PDF). American Museum Novitates. 2002, 3381: 44.
^(英文)徐星、汪筱林、吴肖春.A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China. Nature. 16 September 1999, 401: 262–266.doi:10.1038/45769.
^(英文)Turner, A.H.; Makovicky, P.J.; and Norell, M.A.Feather quill knobs in the dinosaur Velociraptor(pdf). Science. 2007, 317 (5845): 1721.
^(英文)Prum, R.; Brush, A. H. The evolutionary origin and diversification of feathers. The Quarterly Review of Biology. 2002, 77: 261–295.
^Senter, P. Comparison of Forelimb Function Between Deinonychus And Bambiraptor (Theropoda: Dromaeosauridae). Journal of Vertebrate Paleontology. 2006, 26 (4): 897–906. doi:10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2. }
^(英文)Forster, C. A. The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana. The Mosasaur. 1984, 2: 151–163.
^38.038.1(英文)Wedel, M. J.; Cifelli, R. L. Sauroposeidon: Oklahoma’s Native Giant. Oklahoma Geology Notes. 2005, 65: 40–57.
^(英文)Maxwell, W. D.; Ostrom, J.H. Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. Journal of Vertebrate Paleontology. 1995, 15 (4): 707–712. doi:10.1080/02724634.1995.10011256.
^(英文)Brinkman, Daniel L.; Cifelli, Richard L., Czaplewski, Nicholas J. "First Occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptain-Albian) of Oklahoma". Oklahoma Geological Survey. 1998, (164): 27.
^(英文)Roach, B. T.; D. L. Brinkman. A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. Bulletin of the Peabody Museum of Natural History. 2007, 48 (1): 103–138.
^Li, Rihui; Lockley, Martin G.; Makovicky, Peter J.; Matsukawa, Masaki; Norell, Mark A.; Harris, Jerald D.; Liu, Mingwei. Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China. Naturwissenschaften. 2007, 95 (3): 185. doi:10.1007/s00114-007-0310-7.
^43.043.1(英文)Fowler, D.W.; Freedman, E.A.; Scannella, J.B.; Kambic, R.E.The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds. PLoS ONE. 2011, 6 (12): e28964. PMID 22194962. doi:10.1371/journal.pone.0028964.
^(英文)Therrien, F., Henderson, D.M. and Huff, C.B. (2005). "Bite me: biomechanical models of theropod mandibles and implications for feeding behavior." Pp. 179–237 in Carpenter, K. (ed.), The Carnivorous Dinosaurs. Indianapolis: Indiana University Press.
^(英文)Gignac, P.M.; Makovicky, P.J.; Erickson, G.M.; Walsh, R.P. A description of Deinonychus antirrhopus bite marks and estimates of bite force using tooth indentation simulations. Journal of Vertebrate Paleontology. 2010, 30 (4): 1169–1177. doi:10.1080/02724634.2010.483535.
^46.046.146.2(英文)Ostrom, J. H.On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus. Breviora. 1976, 439: 1–21.
^(英文)Li, Rihui; Lockley, Martin G.; Makovicky, Peter J.; Matsukawa, Masaki; Norell, Mark A.; Harris, Jerald D.; Liu, Mingwei. Behavioral and faunal implications of deinonychosaur trackways from China. Naturwissenschaften. 2007, 95 (3): 185–91. PMID 17952398. doi:10.1007/s00114-007-0310-7.
^(英文)Adams, Dawn (1987) "The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs" pg 1–6 in Currie, Philip J. and Koster, E. (eds) Fourth symposium on mesozoic terrestrial ecosystems. Tyrrell Museum, Drumheller, Canada
^(英文)Carpenter, K. Evidence of predatory behavior by carnivorous dinosaurs. Gaia. 1998, 15: 135–144.
^50.050.1Manning, Phil L.; Payne, David; Pennicott, John; Barrett, Paul M.; Ennos, Roland A. Dinosaur killer claws or climbing crampons?. Biology Letters. 2006, 2 (1): 110–112. PMC 1617199. PMID 17148340. doi:10.1098/rsbl.2005.0395.
^51.051.1(英文)Davies, S. J. J. F. Ratites and Tinamous. New York: Oxford University Press. 2002. ISBN 0-19-854996-2.
^(英文)Gilliard, E. T. Living birds of the world. Garden City, NY: Doubleday. 1958.
^(英文)Kofron, Christopher P. Attacks to humans and domestic animals by the southern cassowary (Casuarius casuarius johnsonii) in Queensland, Australia. Journal of Zoology. 1999, 249 (4): 375–381. doi:10.1111/j.1469-7998.1999.tb01206.x.
^(英文)Kofron, Christopher P. Case histories of attacks by the southern cassowary in Queensland. Memoirs of the Queensland Museum. 2003, 49 (1): 335–338.
^(英文)Redford, Kent H.; Peters, Gustav. Notes on the biology and song of the red-legged seriema (cariama cristata). Journal of Field Ornithology. 1986, 57 (4): 261–269. JSTOR 4513154.
^(英文)Carpenter, K. Forelimb biomechanics of nonavian theropod dinosaurs in predation. Senckenbergiana Lethaea. 2002, 82: 59–76. doi:10.1007/BF03043773.
^(英文)Gauthier, J.; Padian, K. Phylogenetic, Functional, And Aerodynamic Analyses Of The Origin Of Birds And Their Flight. (编) M.K. Hecht, J.H. Ostrom, G. Viohl, & P. Wellnhofer (eds). proceedings of the International Archaeopteryx Conference, Eichstätt, 1984. The Beginnings Of Birds. Eischtatt: Freunde des Jura-Museums Eichstätt: 185–197. 1985. isbn 3-9801178-0-4.
^(英文)Senter, P. Comparison of Forelimb Function Between Deinonychus And Bambiraptor (Theropoda: Dromaeosauridae). Journal of Vertebrate Paleontology. 2006, 26 (4): 897–906. doi:10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2.
^(英文)Gishlick, A.D. The function of the manus and forelimb of Deinonychus antirrhopus and its importance for the origin of avian flight. (编) Gauthier, J. and Gall, L.F. New Perspectives on the Origin and Early Evolution of Birds. New Haven: Yale Peabody Museum. 2001: 301–318.
^(英文)Parsons, W.; Parsons, K.Parsons, W. (2003-2005) list of conference proceedings abstracts Morphology And Size Of An Adult Specimen Of Deinonychus antirrhopus, (Saurischia, Theropoda). Journal of Vertebrate Paleontology. 2006, 26 (3): 109A.
^(英文)Parsons, W. L.; and Parsons, K. M.Further descriptions of the osteology of Deinonychus antirrhopus (Saurischia, Theropoda)(PDF). Bulletin of the Buffalo Society of Natural Sciences. 2009, 38: 43–54.
^(英文)Kool, R. The walking speed of dinosaurs from the Peace River Canyon, British Columbia, Canada. Canadian Journal of Earth Science. 1981, 18 (4): 823–825. doi:10.1139/e81-077.
^Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 331-336.
^(英文)Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

相关资料




- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读


知识互答

关于我们

APP下载



{{item.time}} {{item.replyListShow ? '收起' : '展开'}}评论 {{curReplyId == item.id ? '取消回复' : '回复'}}