






翼龙目
发现历史
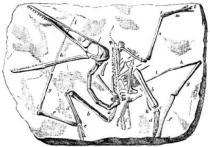
古老翼手龙模式标本的素描,由理查德·莱德克在1888年绘制
第一个翼龙类化石是在1784年由意大利自然学家Cosimo Collini发现。Collini当时将这些化石误认为海生动物,将它们的长前肢充当桨来使用。一些科学家持续支持这个海生动物假设,直到1830年,德国动物学家Johann Georg Wagler仍提出翼手龙属的前肢是用来游泳的。在1801年,乔治·居维叶(Georges Cuvier)研究一个在德国发现的化石,并首次提出这种动物是种飞行动物。在1809年,乔治·居维叶将这个德国化石命名为翼手龙属(Ptero-dactyle)。之后因为科学名称的标准化,翼手龙属的正式属名改成Pterodactylus。
自从1784年在索伦霍芬石灰岩层(年代属于侏罗纪晚期)发现第一个翼龙类化石后,当地沉积层已发现29种翼龙类。在1828年,玛丽·安宁(Mary Anning)在英国莱姆里吉斯发现著名的双型齿翼龙。在1834年,Johann Jakob Kaup建立翼龙目(Pterosauria);然而,在最早期的研究中,有时会采用Ornithosauria(意为“鸟类蜥蜴”)一词。

古魔翼龙的头颅骨,发现于巴西桑塔纳组
大多数翼龙类化石不是保存良好。它们的骨头是中空的,当沉积物堆积在它们身上时,骨头会被压平。目前保存最良好的化石在1974年发现于巴西的Araripe Plateau。当沉积物堆积到这些化石上时,沉积物会压缩骨头,而非压碎骨头。
现在大多数古生物学家认为翼龙类是采动力飞行,而非原先认为的滑翔飞行。翼龙类化石已在北美、南美、英国、欧洲、非洲、亚洲、澳洲等地发现,除了南极洲以外。目前已发现至少60属翼龙类,体型小如小型鸟类,大型翼龙类的翼展可超过10米。
生理结构
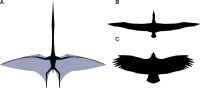
A.风神翼龙、B.漂泊信天翁、C.南美秃鹫的翅膀比较图(不按照比例)

翼龙类、蝙蝠、鸟类的翅膀结构图
翼龙类的生理结构,因为飞行演化的需求,而与它们祖先的生理结构有大幅差异。翼龙类的骨头中空,内有空气,类似现代鸟类的骨头。它们拥有隆脊状胸骨,可让飞行相关肌肉附着在上,还有加大的脑部,显示出与飞行有关的特化特征。在某些较晚的翼龙类里,肩膀部位的脊椎愈合成一节联合背椎(Notarium,一种在鸟类与某些翼龙类肩膀出现的复合脊骨),可在飞行时使身体坚挺,并提供肩胛骨稳定的支撑。
翼
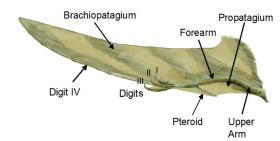
翼龙类的翅膀结构
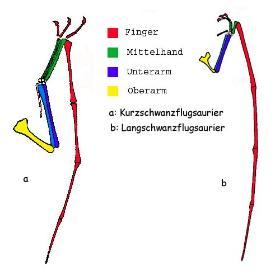
翼龙类的翼部结构(a短尾,b长尾),上臂(黄)、前臂(蓝)、掌骨(绿)、指骨(红)
翼龙类的翼膜由皮肤与其他软组织构成,由不同形式的紧密纤维补强。翼膜连接至极长的第四手指与身体侧面(或是后肢)之间。
过去的观点认为,翼龙类的翼膜构造简单,仅由皮肤构成。但现在的观点认为,翼龙类的翼膜构造相当复杂,具有高度气动性,适合主动飞行。翼膜是由一种排列紧密的强韧纤维所构成,名为“Actinofibrils”。这种翼龙类专有的独特纤维,以十字方式互相交错、重叠。目前无法得知这种独特纤维是由角质、肌肉、或者是具弹性物质所构成,因此也无法得知其实际功能。研究人员推测,这种纤维可能有使翼膜坚韧、或补强的作用。翼膜由皮肤、极薄的肌肉构成,由不同形式的紧密纤维补强,并具有复杂的血管系统。
某些大型标本的翼部骨头内有中空空间,证实某些翼龙类具有类似鸟类的呼吸系统。此外,根据至少一个标本的软组织,显示其呼吸系统延伸到翼膜内部。
翼龙类的翼膜可以分成三个部分。第一个部分是前膜(Propatagium),连接手腕部到肩膀,位于翼膜最前端,是飞行时首先遭遇到空气的部分。某些化石证据显示,前三根手指之间也连接者前膜。翼膜的主要部分是臂膜(Brachiopatagium),从极长第四指延伸至身体两侧(或后肢)。但臂膜连接至身体两侧的哪个位置、是否连接至后肢,仍有争议。某些翼龙类的后肢之间连接者膜,可能延伸至尾巴,称为尾膜(Uropatagium)。根据索德斯龙的化石,其尾膜仅连接后肢,并没有连结至尾巴。研究人员多认为,喙嘴翼龙类具有宽广的尾膜,而翼手龙类的尾膜仅连接者后肢之间;无齿翼龙是个例外,其尾膜连接者后肢、尾巴。
从手腕延伸出的翅骨(Pteroid),是一种翼龙类专有的骨头,协助支撑手腕部到肩膀的前膜。化石证据显示,前三根手指之间可能也连接者翼膜。因此,前膜的面积可能更大。古生物学界对翅骨角度、位置的看法相当不同。大卫·安文等人认为,翅骨的角度往前,扩大前膜的面积。但论点这是非常有争议的。在2007年,克里斯·班尼特认为翅骨的角度朝向身体,而非朝前,如同传统的看法。根据组织学、个体生长学、形态学等层面的证据,皆显示翅骨本身是个骨头,而非骨化的软骨。古生物学界对于翅骨的起源还没有定论,目前有:从腕骨延伸出、第一掌骨延伸而出、或是新演化的骨头…等说法。
翼龙类的手腕有两块近侧腕骨、四块末侧腕骨(不计算翅骨的状况),显示它们的手腕经过明显特化过程。在成年个体标本,这两块近侧腕骨愈合成一块,而其中三块末侧腕骨愈合成一块。唯一未愈合的腕骨位于手腕中间,位于末侧腕骨、翅骨之间,连接至末侧腕骨的垂直突出面,另一侧则是以凹处连接翅骨。
古生物学家们对于臂膜是否也连接到后肢,连接至后肢的哪个部位,目前还没有定论。喙嘴翼龙类的索德斯龙、蛙嘴龙科的热河翼龙、还有桑塔纳岩层发现的某种翼手龙类化石,证实至少某些物种的翼膜有连接至后肢。然而,现代蝙蝠与飞鼠翼膜的连接方式上有相当大的不同,而不同种类的翼龙类可能也有不同的翼膜连接方式。研究显示翼龙类的四肢比例有相当大的不同,可能反映了不同的翼展方式。
许多翼龙类有蹼状脚掌,可能不是全部都有,蹼状脚掌可能具有气动上的作用,但也有研究认为蹼状脚掌是种游泳的证据。某些现代滑翔动物也具有蹼状脚掌,例如鼯猴。
颅骨、牙齿

一颗翼龙类的牙齿,可能属于科罗拉多斯翼龙
大部分翼龙类具有修长的口鼻部。许多翼龙类的嘴部布满针状牙齿;而某些衍化物种则没有牙齿,具有类似现代鸟类的狭长喙状嘴,例如无齿翼龙科、神龙翼龙科。某些标本的喙状嘴则保存了角质组织。
大部分主龙类的眼部前方有数个洞孔;而翼龙类的鼻孔与眶前孔连接,形成一个鼻眶前孔(Nasoantorbital fenestra)的大型洞孔。鼻眶前孔可能具有减轻重量功能,有助于飞行。
头冠
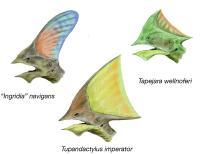
三种古神翼龙科的头冠,分别为:T. navigans(左上)、沃氏古神翼龙(右上)、雷神翼龙(下)。
许多翼龙类的头顶具有头冠。第一个被发现头冠的翼龙类是无齿翼龙,其尖形头冠朝向头后方。某些物种的头冠形状特殊、巨大,由骨质分叉支撑者角质组织所构成,例如古神翼龙科与夜翼龙。
自从1990年代以来的新发现化石与详细研究,发现翼龙类普遍具有头冠,这与过去的观点不同。翼龙类的头冠主要由角质组织构成,甚至是全部,因此很少在化石化过程中保存下来。科学家借由紫外线摄像技术,检验出翼手喙龙与翼手龙属的化石也具有头冠,而过去没有被发现。根据过去的理论,只有衍化的翼手龙类具有头冠;而近年发现的翼手喙龙、奥地利翼龙也具有头冠,证实部分喙嘴翼龙类也具有头冠。

索德斯龙的想像图,后肢之间连接者尾膜,但没有连接至尾巴
毛
翼龙类并没有发现羽毛证据,但至少部分翼龙类覆盖者类似毛的丝状结构,类似哺乳类的毛发,但非同源演化的结果。翼龙类的毛不同于哺乳类的毛发,而是独特的结构,两者为趋同演化的后果。虽然在有些研究里,翼膜上的纤维被误认为毛,而有些化石的头部与身体上的确有丝状结构物痕迹,例如索德斯龙、热河翼龙。翼龙类的毛不类似现今蝙蝠的毛发,这是另一个趋同演化的例子。翼龙类演化出毛,是基于飞行的需求,也显示翼龙类是温血动物。
在2009年的热河翼龙研究,古生物学家亚历山大·克尔纳将翼龙类的毛建立一个专有名词,“Pycnofibre”,意为密集的丝状物。
古生物学
飞行
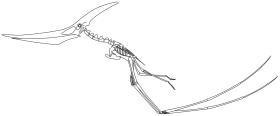
刚弹射起飞姿势的无齿翼龙
关于翼龙类的飞行模式、双翼运动方式、以及起飞方法,古生物学界还没有定论。在2008年,日本东京大学研究人员佐藤克文认为翼龙类无法飞行。佐藤克文在印度洋南部的克罗泽群岛进行特殊研究,目的在测试大型海鸟的飞行能力。他将三号电池大小的加速度计贴在五个不同种类、二十八只鸟的羽翼上,其中包括世界最大的飞鸟信天翁。根据佐藤克文的计算,重量超过四十公斤的鸟在无风状态下,翅膀拍动速度无法达到足以飞翔的速度。信天翁的重量为二十二公斤,还是能够飞行。依据这个基准,大型翼龙无法翱翔高空。不过,研究报告指出,比较这两种飞行动物时,还必须考虑它们在解剖结构、生理学与环境上的差异。在书籍《Posture, Locomotion, and Paleoecology of Pterosaurs》里,则认为白垩纪晚期的大气层的氧气含量高,翼龙类可以在这种环境中飞行。但是,上述两者都推测翼龙类以类似鸟类的方式飞行,而且无法将某些巨大翼龙类的体型列入计算(例如神龙翼龙科、古神翼龙科)。德恩·奈许(Darren Naish)则认为中生代与现代的大气层组成不同,并非大型翼龙类能够飞行的原因。
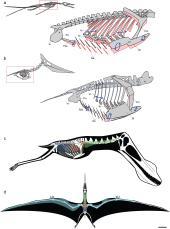
当翼龙类呼吸时,身体骨骼、气囊系统的移动
朴茨茅斯大学与约翰·霍普金斯大学的古生物学家马克·维顿、麦克·哈比卜提出翼龙类的起飞方式假设,当翼龙类起飞时,会将身体呈预备状态,再用四肢力量同时推动身体向前,进而展开双翼飞行,类似弹射器的弹射方式。当翼龙类在天空飞行时,时速可达每小时120公里,一次可飞行数公里。
在1985年,史密森尼学会委托航空工程师保罗·麦卡克莱迪(Paul MacCready)进行翼龙类的飞行模式测试,他制造一个1/2大小的风神翼龙模型,并以弹射装置进行多次测试,并把过程拍摄成IMAX电影《On the Wing》。但这个模型有一些生理结构上的错误,例如尾巴装设垂直、水平方向的控制舵,以协助模型起飞,但翼龙类的尾巴没有这些额外组织。另外,由于模型尾巴过长,无法实际呈现翼龙类重心分布对飞行的影响。
气囊与呼吸系统
在2009年,一份研究显示翼龙类的骨骼具有充满空气的空间,应该内有气囊,类似现代鸟类的呼吸系统,可使翼龙类的呼吸更有效率、并减轻它们的体重,可协助翼龙类的飞行。
神经系统
因为翼龙类的头颅骨非常易碎,研究它们的头颅骨是非常困难的。但在2003年,俄亥俄大学的劳伦斯·威特默(Lawrence Witmer)等人使用X光电脑断层扫描,扫描明氏喙嘴翼龙(Rhamphorhynchus muensteri)、桑塔纳古魔翼龙(Anhanguera santanae)的脑部,建立起两个物种的脑部3D影像,并发现这两个物种的小脑有巨大的绒球(Flocculus)。绒球是用来整合来自关节、肌肉、皮肤、与平衡器官的讯息。
翼龙类的绒球占了它们脑部体积的7.5%,比其他脊椎动物还大。鸟类的绒球也比其他动物大,但只占脑部的1%~2%。
绒球传送神经讯号,让眼部肌肉产生小而自动的运动。这让动物的视网膜产生稳定的影像。翼龙类有大型绒球的原因,可能是因为它们巨大的翼,巨大的翼意味者有大量的感应讯息要处理。
地上运动
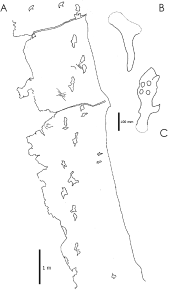
发现于法国的足迹化石,可能属于神龙翼龙科
翼龙类的臀窝是稍微朝向侧上方,股骨头适度的往内侧弯,显示翼龙类是半直立步态。当它们飞行时,大腿可能抬高到与身体平行,类似现代的滑翔蜥蜴。
过去的古生物学家,对于翼龙类在地上移动时是采四足方式或二足方式有重大争论。在1980年代,古动物学家凯文·帕迪恩(Kevin Padian)指出小型翼龙类有较长的后肢,例如双型齿翼龙,它们除了飞行以外,在地面行走与奔跑时可能采二足方式,如同现代的走鹃。现在已发现大量的翼龙类足迹化石,呈现出明显的后肢四趾、前肢三趾的足迹,可证实翼龙类在地上移动是采取四足方式移动较大型翼龙类有较小的后肢与大型的身体前半段,一般认为它们在地面上移动时使用四肢。根据目前所发现的翼龙类足迹化石,可以发现它们正在涉水或找食物,还没有发现可证明它们在飞行或滑翔的足迹化石。

足迹化石显示,诺氏风神翼龙是采四足方式行走

以四足方式觅食的寇氏翼手龙
大部分的脊椎动物是趾行动物,行走时以脚趾接触地面,脚踝不接触地面;从足迹化石显示,翼龙类行走时以脚掌接触地面,类似人类与熊,都属于跖行动物。神龙翼龙科的足迹化石显示,至少有部分翼龙类行走时采取后肢直立步态,而非往后肢两侧延展的步态。
传统的观念认为,翼龙类在地面行动时相当笨拙、不方便;但近年研究发现,至少部分翼龙类(尤其是翼手龙亚目)能够顺利的行走、奔跑。与其他翼龙类相比,神龙翼龙科与鸟掌翼龙科的前肢相当长;神龙翼龙科的手臂骨头与掌骨特别地修长,它们的前肢比例,接近善于奔跑的有蹄类哺乳动物。它们的后肢不适合高速奔跑,但步伐比其他翼龙类更大。神龙翼龙科可能无法奔跑,但它们可以快速、有效率地行走。
借由比较翼龙类与现代鸟类的手掌、脚掌与身体的比例,科学家可以推测翼龙类在地表的生活方式。与神龙翼龙科的体型与后肢相比,它们的脚掌相当小,脚掌长度是胫骨的25%到30%。这显示神龙翼龙科较适合在干燥、硬的地面上行走。无齿翼龙的脚掌较长,长度是胫骨的47%。滤食性的翼龙类(例如梳颌翼龙超科)具有非常大的脚掌,举例而言,翼手龙属的脚掌长度是胫骨的69%,而南翼龙的脚掌长度是胫骨的84%,大的脚掌/胫骨比例代表它们适合在软而泥泞的地面行走,类似现代鸟类。
掠食
过去曾发现棘龙科恐龙以翼龙类为食的化石证据。在《自然》(Nature)杂志的2004年7月版,古生物学家埃瑞克·比弗托(Eric Buffetaut)提出在一个早白垩纪的翼龙类化石的三个颈椎上,发现了棘龙科恐龙的断裂牙齿。由于这些脊椎骨的关节仍连结在一起,所以这只翼龙类未被吞食并消化。
繁衍

发现于索伦霍芬石灰岩的幼年个体标本
关于翼龙类的繁衍行为的直接、间接证据很少。在中国辽宁省的一个采石场发现了一个翼龙类的蛋,同一个挖掘地点也发现了许多著名的有羽毛恐龙。这个蛋被压扁,但没有破碎的迹象,显示这个蛋有皮革质的外壳,如同今日的蜥蜴。胚胎的翼膜已经发展良好,显示翼龙类出生后不久就可以飞行。
在2011年发现的一个达尔文翼龙化石,后肢之间有一个蛋化石,也是质地软的革质蛋,如同现代爬行动物;而现代鸟类则是质地较硬的钙质蛋。与产卵的母体相比,这颗蛋化石非常小。在2007年,一个关于翼龙类蛋壳结构与组成的研究,指出它们可能会将蛋掩埋到土壤中,类似现代鳄鱼与乌龟。对于早期的翼龙类,将蛋掩埋到土壤孵化,可以减轻母体本身的重量,但会限制翼龙类所能生存的地理环境;在鸟类出现后,更会面对鸟类竞争生存环境,而面临劣势。另一种可能则是将蛋置于身体之下,直到孵化前,类似某些蜥蜴的作法,但大部分主龙类不采用此方法。
目前已发现数个刚孵化不久的幼年个体,包含翼手龙科、喙嘴翼龙科、梳颌翼龙科、与神龙翼龙科。这些幼年个体的骨头已呈现高度的硬化,双翼比例已接近成年个体。在翼龙类的早期研究历史,幼年个体、成年个体常被归类于相同属的不同种。德国索伦霍芬石灰岩、巴西的海相沉积层可同时发现成年个体、非常年轻个体,例如翼手龙属、喙嘴翼龙、巴西的南翼龙,可以证实这些个体可能飞跃潟湖的中央时,摔落并淹死。这也显示这些幼年个体已具有飞行能力。

飞越海面的无齿翼龙
目前不确定翼龙类是否有亲代养育行为,但根据它们相当年轻就有飞行能力、海相沉积层的成年与幼年个体化石,显示幼年个体并非完全依靠亲代。或者是只经过短暂的亲代养育后,当双翼已具有飞行能力时,幼年个体就会离巢生活。另一种可能是,它们只借由原本从蛋黄吸收的营养来渡过刚孵化后几天,之后开始独自觅食,完全不靠亲代养育。
翼龙类的不同类群,具有不同的生长模式。早期、原始的喙嘴翼龙类,以喙嘴翼龙为例,刚孵化第一年的平均生长率为130%到173%,略快于现代短吻鳄。当喙嘴翼龙达到性成熟后,成长速率就会缓慢,之后再经过三年才达到最大体型。晚期、进阶型的翼手龙类,以无齿翼龙为例,刚孵化第一年就成长到成年个体体型。翼手龙类的生长模式为有限生长,当成长到完全体型后,就不会继续成长。
活跃时间
在2011年,科学家比较数种翼龙类、现代鸟类与爬行动物的巩膜环大小,以得出它们的作息与活跃时间。翼手龙属、掘颌龙、妖精翼龙属于日行性动物,梳颌翼龙、南翼龙、喙嘴翼龙则是夜行性动物。古神翼龙属于无定时活跃性的动物,觅食、移动行为跟白天黑夜没有正相关,只休息短暂时间。根据这个研究结果,梳颌翼龙、喙嘴翼龙的生活方式可能类似现代夜行性海鸟,在夜间捕食鱼类;南翼龙的生活方式可能类似某些雁形目,在夜晚采滤食方式以水中的小型动物为食。德国索伦霍芬石灰岩同时发现梳颌翼龙、喙嘴翼龙、掘颌龙、翼手龙属,它们可能占据不同生态位、有不同生活方式与食物来源。
演化与灭绝
起源

生存于三叠纪晚期的斯克列罗龙,可能是翼龙类祖先的近亲
翼龙类的骨骼结构因为适应飞行而有大幅改变、特化,而且目前没有它们最直接祖先的化石证据,所以目前对翼龙目的演化起源了解不多。因为它们的踝部结构,翼龙类被认为与恐龙是近亲。目前已有数个相关理论,其中近年最盛行的理论是类似斯克列罗龙(Scleromochlus)的鸟颈类主龙、能以二足方式站立的原始主龙形类派克鳄、或者是类似沙洛维龙的原蜥形目动物。目前至少有一位翼龙类专家,大卫·安文(David Unwin),认为这些动物因为个别的生理特征,都不符合翼龙类祖先的假设。
克里斯·班尼特、大卫·彼得斯曾先后提出翼龙类属于原蜥形目,或者是原蜥形目的近亲。大卫·彼得斯利用电脑绘图软件,企图找出原蜥形目、翼龙目的关联。克里斯·班尼特在试图找寻翼龙类、恐龙的趋同演化特征时,发现翼龙类除去后肢特征时,相当类似原蜥形目,因此提出原蜥形目、翼龙类有演化关系。在2007年,麦可·班顿(Michael Benton)等人提出不同看法,认为翼龙类与恐龙是近亲,即使不考虑后肢特征,翼龙类仍非常类似恐龙。他们并批评大卫·彼得斯的研究,不是研究原始化石与资料,而采用具争议性的分析途径,最后导论出原蜥形目、翼龙类是近亲。。大卫·彼得斯则坚持电脑绘图软件,可用在化石研究上。班顿等人认为虽然目前没有翼龙类最直系祖先的化石证据,但翼龙类很明显是属于主龙类,尤其是鸟颈类主龙演化支。根据他们的分析,翼龙类是斯克列罗龙的姊妹分类单元,或者是从介于斯克列罗龙与兔鳄之间的物种演化而来。在2011年,斯特林·奈斯比的早期主龙类研究,也支持翼龙类、斯克列罗龙互为姊妹分类单元的理论。
因为翼龙类被证实没有树栖生活的演化适应,所以它们的飞行演化途径被认为跟鸟类不同路线,鸟类的飞行演化途径是“从树往下的”。大多数方案认为翼龙类从长腿的陆地奔跑动物演化而来,如斯克列罗龙或沙洛维龙,上述两者都有皮膜,从后腿延展至身体或尾巴。这些研究显示翼龙类的飞行演化途径是“从地面往上”,或是攀爬悬崖。在2008年,一份研究显示最早的翼龙类是群树栖、食虫的小型动物。
种系发生学与分类
在古生物学研究历史中,翼龙目的科学分类是很困难的,因为它们的化石纪录有许多间断。许多新发现化石填补了这些断层。传统上,翼龙目被分为两个亚目:
喙嘴翼龙亚目(Plieninger, 1901):一群早期、原始翼龙类,许多物种有长尾巴,双翼有短指骨。它们体型小,手指仍然适合攀爬。喙嘴翼龙类出现在晚三叠纪,持续存活到晚侏罗纪。喙嘴翼龙类是并系群,其中一群演化出晚期的翼手龙类,而非两者有共同祖先;所以随者亲缘分支分类法研究的盛行,本名词逐渐很少在科学文献现。
翼手龙亚目(Plieninger, 1901):更多样性的翼龙类,有短尾巴,双翼有长指骨。翼手龙类出现在中侏罗纪,在白垩纪-第三纪灭绝事件中灭绝。
以下列表是根据大卫·安文(David Unwin)在2006年的研究:

喙嘴翼龙生存于晚侏罗纪,属于喙嘴翼龙亚目
浙江翼龙生存于白垩纪的中国,属于神龙翼龙科
翼龙目 Pterosauria
翼龙类各演化支的彼此关系并未确定。然而几个较新的研究开始让它们更明确。以下演化树是简略参考大卫·安文的研究:
灭绝
传统理论认为,早期鸟类的竞争,导致许多翼龙类灭绝。到了白垩纪末期,只有发现大型翼龙类;而小型翼龙类已消失,生态位被早期鸟类取代。但是,化石纪录缺乏小型翼龙的现象,也可能是因为它们的骨架脆弱、难以保存所致,而跟鸟类的生物多样性无关。白垩纪末灭绝事件灭绝了恐龙、翼龙类、与许多其他动物。另一种解释是,大多数翼龙类发展成依靠海洋的生活模式。所以当白垩纪末灭绝事件严重地影响翼龙类赖以为生的海洋动物时,翼龙类跟者灭绝。但是,神龙翼龙科与帆翼龙科是栖息于内陆,并非依靠海洋维生。
近年发现的数个新物种,生存于坎潘阶/马斯特里赫特阶,例如:鸟掌翼龙科、无齿翼龙科、夜翼龙科、古神翼龙科。这些新发现物种,显示白垩纪晚期的翼龙类仍保持多样性,相对于白垩纪早期,只是小幅度衰退,不同于过去理论。
著名属
大众文化

风神翼龙的模型,因为英国皇家学会300周年而架设于伦敦市区
如同其近亲恐龙,翼龙类是大众文化中的一个常见主题。在大众媒体的恐龙形象,常随者古生物学界的研究而有新改变;但是翼龙类的大众形象,仍多停留在20世纪中期的过时研究。
大众媒体、一般民众常使用翼手龙(Pterodactyls)来称呼这些中生代飞行爬行动物,这名词所形容的动物外表经常是无齿翼龙、喙嘴翼龙,或是数个翼龙类的幻想混合体。许多儿童的玩具与卡通节目,将翼手龙(Pterodactyls)描绘成具有类似无齿翼龙的冠饰,加上类似喙嘴翼龙的长尾巴与牙齿,但这种幻想生物从未实际存在过。然而,ˊ至少有一种翼龙类的确有无齿翼龙的冠饰,同时具有牙齿;玩具翼龙(Ludodactylus)的学名字意思为“玩具翼手龙”,因为它们外表类似儿童的翼手龙玩具外貌。
这种幻想、混合体翼手龙,还出现在阿瑟·柯南·道尔(Arthur Conan Doyle)的小说《失落的世界》(The Lost World)与其改编电影、以及1933年的电影《金刚》(King Kong)。在1966年电影《公元前一百万年》(One Million Years B.C.),早期电影特效专家雷·哈利豪森把翼手龙的双翼错误塑造成蝙蝠的多指双翼。雷·哈利豪森宣称,当他用实体模型拍摄时,为了增强双翼的结构,而采用蝙蝠的双翼结构。不过当时大众媒体的翼手龙形象,常被建立成这种类似蝙蝠的双翼。在2001年的特效电影《侏罗纪公园Ⅲ》(Jurassic Park III),出现一只无齿翼龙。古生物学家戴维·霍尼(Dave Hone)发现该电影仍保有过时、错误的翼龙类形象,例如:电影里的无齿翼龙仍保有牙齿、错误的筑巢方式、双翼应该是由肌肉与紧密纤维所构成。《哥斯拉》系列电影中的飞行怪兽拉顿(ラドン),其名称与形象来自于翼龙类。
活生翼龙恶作剧
在伦敦新闻画报1856年2月9日的第166页里,报导宣称在法国的圣迪济耶到南锡之间的地铁隧道,一群工人挖掘到侏罗纪石灰岩层时,一只大型动物从里面跌了出来。它拍动了它的翅膀,叫了几声就死了。根据工人的说法,这个动物翼展长达10呎,四肢都附有膜,黑色皮革般皮肤,嘴巴里有牙齿,脚掌上有利爪。一个当地古生物学学生将这动物鉴定为翼手龙(Pterodactyl)。
这个故事是个恶作剧,导因于法国与普鲁士在古生物学上的竞争。巴伐利亚州的索伦霍芬石灰岩层发现了许多有价值的化石,包括始祖鸟,都被德国古生物学家骄傲地发表。故事中隧道是挖穿与索伦霍芬石灰岩层年代相似的石灰岩层,故事背后隐藏了法国的竞争心态。
备注与参考资料
(英文) Unwin, David M. (2006). Pterosaurs From Deep Time. Pi Press: New York. ISBN 978-0-13-146308-0
(英文) Wellnhofer P (1991): Illustrated Encyclopedia of Pterosaurs, Crescent Books
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

相关资料





- 有价值
- 一般般
- 没价值















推荐阅读


关于我们

APP下载


