






二叠纪-三叠纪灭绝事件
年代测定
在西元二十世纪之前,二叠纪与三叠纪交界的地层很少被发现,因此科学家们很难准确地估算灭绝事件的年代与经历时间,以及影响的地理范围 。在1998年,科学家研究浙江省长兴县煤山附近的二叠纪/三叠纪岩层,他们采用铀-铅测年方法,研究锆石中的铀/铅比例 ,估计二叠纪-三叠纪灭绝事件的发生年代为2亿5140万年前(误差值为30万年),并发现此后有随者时间持续增高的灭绝比例 。在灭绝事件发生时,全球各地的碳13/碳12比例极速下降约9‰ 。因为二叠纪/三叠纪的界线难以用放射性定年法测定,科学家们多用急遽下降的碳13/碳12比例,测定岩层中的二叠纪/三叠纪交界 。
在二叠纪与三叠纪交界时期,陆地与海生真菌曾有急遽的增加,原因应是当时有大量的植物与动物死亡,使得真菌得以迅速繁衍 。因为二叠纪/三叠纪的界线难以用放射性定年法测定,也缺乏参考用的标准化石,古动物学界一度使用真菌的繁衍高峰,测定岩层中的二叠纪/三叠纪交界。但是真菌定年法的最初提倡者,指出真菌的繁衍高峰,可能是三叠纪最早期生态系统的反复发生现象 。真菌的繁衍高峰理论在最近几年遭到许多质疑,例如:最常发现的真菌孢是 Reduviasporonites ,在近年被发现其实是藻类化石、 、真菌的繁衍高峰似乎不是全球性现象 、在许多地层中,真菌的繁衍高峰与二叠纪/三叠纪界线并不一致 。藻类被误认为真菌胞,可能代表三叠纪时期全球各地湖泊增加,而非生物的大量死亡与腐烂 。在2009年的一个研究,则以生物化学方式分析 Reduviasporonites 的组成,例如:碳同位素、氮同位素、碳/氮比例,发现 Reduviasporonites 的组成接近真菌 。
关于这次灭绝的过程、经历时间、以及不同生物群的灭绝模式,仍然相当模糊。一些证据显示这次灭绝事件持续约数百万年,而在二叠纪的最后100万年,急遽地达到高峰 。在2000年,科学家研究浙江煤山的许多富含化石的地层,借由统计分析指出,这次灭绝事件只有一个灭亡高峰期 。但是,之后近年的研究指出不同的生物群在不同的时间点灭亡;举例而言,虽然年代难以确定,介形纲(Ostracoda)与腕足动物的大规模灭绝,先后相差了72万年到122万年之间 。在格陵兰东部的一个化石保存良好的地层,动物群的衰退集中在6万到1万年之间,而植物群的衰退则持续约数十万年之久,但也显示灭绝事件造成全面性的影响 。一个较早期的研究,认为当时有两个灭绝高峰期,相隔500万年,这段期间的生物灭绝高于正常标准;而最后的灭绝高峰,造成当时约80%的海洋生物灭亡,其余海生生物则多在第一次的灭绝高峰与灭绝间期消失;近年仍有研究引用这个假设 。若根据此一理论,第一次的灭绝高峰发生于二叠纪瓜达鹿白阶末期 。
瓜达鹿白阶 (法语) 末期灭绝事件对海生生物的冲击,随者地区与物种的不同,而有不同程度的影响。举例而言,腕足动物与珊瑚有严重的大量灭亡 。所有的恐头兽类灭亡,仅有一属继续存活 ;在有孔虫门中,体型大型的纺锤虫目(Fusulinida)也仅有费伯克蜓科(Verbeekinidae)继续存活 。
灭绝模式
海中生物
在二叠纪-三叠纪灭绝事件中,海生无脊椎动物的灭亡程度最大。在中国南部的一些富含化石的地层,在329属的海生无脊椎动物中,有280属在二叠纪最后两个沉积层(以发现牙形石化石为标准)消失 。
根据统计分析的结果,二叠纪末海洋生物的物种数量降低,多是因为灭亡,而非物种形成的减少 。受到此次灭绝事件影响最严重的生物群,是具有碳酸钙外壳的海洋无脊椎动物,尤其是吸收环境中二氧化碳以制造外壳的物种 。
在正常时期,底栖生物的灭亡率高。这次灭绝事件使底栖生物的灭亡率,达到正常时期的数倍 。以灭绝比例来看,海洋生物遭受灾难性的灭亡 。
继续存活的海生无脊椎动物则包含:腕足动物门(它们的数量在灭绝事件后持续了一段缓慢的衰退)、菊石亚纲的齿菊石目与海百合纲(几乎灭亡,但后来再度繁盛、多样化)。
存活比例最高的海洋生物,通常具有良好的循环系统、复杂的气体交换机制、以及较少的钙化组织。而钙化组织最多的物种,则是灭亡比例最高的生物 。以腕足动物为例,继续存活的物种体型较小,且是多样化生物群落中的数量稀少成员 。
自从二叠纪中期 罗德阶 ( 英语 : Roadian ) 之后,菊石亚纲已经历长时间的衰退,达约3000万年之久。瓜达鹿白阶(Guadalupian)末期的有限度灭绝事件,使菊石的差异性大幅减少,环境因素可能导致这次的灭绝事件。菊石的低多样性与低差异性持续到二叠纪末灭绝事件;然而,二叠纪末灭绝事件的范围广泛、过程迅速。在三叠纪,菊石开始迅速多样化,但物种之间的差异性仍然不大 。
在二叠纪时期,菊石的形态空间逐渐有限。在进入三叠纪的数百万年后,菊石再度发展出原始的形态空间,但不同演化支已出现差异 。
陆地无脊椎动物
在二叠纪时期,陆地的昆虫与其他无脊椎动物非常繁盛、多样化,其中也包含地表上曾出现过的最大型昆虫。发生在二叠纪末的灭绝事件,是目前已知惟一的昆虫大规模灭绝事件 ,其中有八、九个昆虫的目消失,而至少十个目的物种减少。古网翅总目(Palaeodictyopteroidea)在二叠纪中期开始衰退,这段时间可能与植物群的变化有关系。昆虫的衰退高峰发生于二叠纪晚期,这段时间的高峰则可能与植物群的变化(由气候变化造成)无直接关联 。
在二叠纪-三叠纪灭绝事件的前后,昆虫的化石有极大的差异。许多古生代的昆虫多在二叠纪-三叠纪灭绝事件后消失,只有舌鞘目、Miomoptera(内翅总目的一目)、原直翅目继续存活到中生代。外翅总目、单尾目、古网翅总目、Protelytroptera(舌鞘目的一亚目)、原蜻蜓目在二叠纪末灭亡。在一些保存状态良好的三叠纪晚期地层中,通常可发现与现代昆虫族群相关的化石 。
陆地植物
植物生态系统
陆地植物的化石纪录很少见,大多数是花粉与孢子。所有大型灭绝事件对植物造成的影响,只有到科的层级 。若以种层级来看,减少的物种大部分可能是埋藏过程造成的假象 。若以优势物种与地理分布来看,可发现植物生态系统有重大的变动 。
在二叠纪/三叠纪交界时,优势的植物群改变,许多陆地植物突然开始衰退,例如荷达树(裸子植物)与舌羊齿(种子蕨) 。在二叠纪/三叠纪交界之后,原本占优势的裸子植物被石松门取代 。
一个针对格陵兰东部沉积层中的花粉与孢子研究,显示在灭绝事件发生前,当地有密集的裸子植物森林。在海生无脊椎动物群衰退时,这个大型森林也开始消失,而小型草本植物开始多样化(包含石松门的卷柏与水韭)。裸子植物后来一度兴盛,然后大量灭亡。在二叠纪末灭绝事件期间与之后,植物群发生数次兴衰循环。优势植物群从大型木本植物变动到草本植物,显示当时许多的大型植物遭到环境改变的压力。植物群落的接续与灭绝,发生于碳13/碳12比例极速下降之后,但两者时间并不一致 。裸子植物森林的复原,花了约400万到500万年 。
煤层缺口
在三叠纪早期的地层中,没有发现煤矿床,而三叠纪中期的地层中,煤矿床薄,而且品质低 。关于煤层缺口的成因有许多假设。有科学家认为,当时有许多新的真菌、昆虫、以及脊椎动物出现,这些具侵略性的动物造成植物的大量死亡。但是,这些动物也因二叠纪-三叠纪灭绝事件而大量灭亡,不太可能是煤层缺口的主因 。有可能所有可形成煤矿的植物,在这次灭绝事件中几乎灭亡;而在1000万年之后,新的植物才适应潮湿、酸性的泥炭沼泽,而出现可形成煤矿的新植物 。非生物性因素,则包含:减少的降雨、或增加的碎屑沉积物 。最后,三叠纪早期的各种沉积层很少被发现,煤层的缺口可能反映了沉积层的稀少状况。有可能形成煤矿的三叠纪早期生态系统,目前还没有被发现 。例如在澳洲东部,当时可能长时间处在寒冷的气候,这种天气状况适合泥炭的形成。在可以形成泥炭的植物中,有95%在这次灭绝事件中消失 。但是,澳洲与南极洲的煤矿床在P-Tr事件发生前,就已经消失 。
陆地脊椎动物
在二叠纪末期,许多陆地脊椎动物消失,而某些演化支甚至几乎灭亡。某些存活的族系,并未长时间继续存在,而其他族系则开始多样化,并长期存活着。有充足证据显示,超过⅔的陆生两栖动物、蜥形纲、兽孔目的科在这个灭绝事件中消失。大型的草食性动物遭受严重的打击。除了前棱蜥科,几乎所有的二叠纪无孔亚纲动物灭亡;龟鳖目具有无孔类的头骨,被认为可能是从前棱蜥科的近亲演化而来(或是双孔亚纲)。盘龙目早在二叠纪末期前就已灭亡。由于二叠纪的双孔亚纲化石很少,无法确定灭绝事件对双孔亚纲所造成的影响;双孔亚纲包含大部分的爬行动物,例如蜥蜴、蛇、鳄鱼、恐龙、以及恐龙的后代鸟类 。
灭绝模式的可能解释
灭亡程度最严重的海生动物,包含:可以制造钙质外壳(例如碳酸钙)、代谢率低、以及呼吸系统较弱的物种,尤其是钙质海绵类、四射珊瑚、床板珊瑚、具钙质外壳的腕足动物、苔藓动物门、棘皮动物门,上述动物中约有81%的属灭亡。而无法制造钙质外壳的近亲,受到的影响较小,例如海葵(演化出现代的珊瑚)。具有高代谢率、呼吸系统良好、以及缺乏钙质外壳的动物,灭亡的幅度较小;牙形石是个例外,约有33%的属消失 。
这种灭绝模式,与组织缺氧产生的效应相符合。在当时,几乎所有的大陆棚海域都有严重的缺氧现象,但缺氧现象无法解释有限度的灭绝模式;因此缺氧不是造成海生生物灭绝的唯一成因。一个针对二叠纪晚期到三叠纪早期大气层的数学模型,指出在这段期间,大气层中的氧气有明显、长时期的衰退;这个衰退不因二叠纪/三叠纪交界而增加,并在三叠纪早期达到最低点,但仍高于现今的比例。由此得知,氧气比例的衰退与灭绝模式并不一致 。
另一方面,海生生物的灭绝模式也与高碳酸血症(二氧化碳过高)产生的效应相符合。当二氧化碳浓度高于正常时,将对生物造成伤害:降低呼吸色素携带氧到组织的能力、使体液更为酸性、妨碍动物形成碳酸钙外壳;当浓度更高时,会使生物进入昏迷状态。除了上述反应,二氧化碳浓度高,会使生物制造钙质外壳更加困难。与陆地生物相比,海洋生物对于二氧化碳浓度的变化相当敏感。溶解于水中的二氧化碳,是氧的28倍。对于进行呼吸作用的动物,二氧化碳的排除,与气体经过呼吸系统膜相关;所以海洋动物会将身体内的二氧化碳保持在低浓度,低于陆地动物。若二氧化碳浓度增加,会妨碍海洋动物合成蛋白质、降低受精概率、形成不完整的钙质外壳 。
目前很少发现跨越二叠纪与三叠纪交界的陆相地层,因此难以详细研究陆地生物的灭绝/存活比例。二叠纪与三叠纪的昆虫有显著的不同,但是在二叠纪晚期到三叠纪早期,有个约1500万年的昆虫化石断层。目前最足以研究二叠纪/三叠纪陆地脊椎动物变化的地层,位于南非的卡鲁盆地(Karoo);但目前仍没有足够的研究可供结论 。
生态系统的复原
早期的研究认为在二叠纪末灭绝事件后,地表的生物迅速的复原,但这些研究多是针对在灭绝存活的先驱生物(Pioneer organisms)的研究结果,例如水龙兽。近年的大部分研究认为,当时经历相当长的时间,生态系统才恢复以往的高度多样性、复杂的食物链、与众多生态位。在灭绝事件后,发生数次的灭绝波动,限制栖息地的复原,因此生态系统花了长时期才完全复原。近年的研究指出,地表生态系统直到进入三叠纪中期,约是灭绝事件后400万到600万年,才开始复原 。某些研究人员更指出,直到灭绝事件后3000万年,进入三叠纪晚期,地表生态系统才完全复原 。

怀马卡里里河,分布于新西兰南岛,属于辫状河
在三叠纪早期,约是灭绝事件后400万到600万年,陆地的植物数量不足,造成煤层缺口,使当时的草食性动物面临食物短缺 。南非卡鲁盆地的河流,从曲折河改变为辫状河,显示该地区的植被曾长时间非常稀少 。
三叠纪早期的主要生态系统,无论是植物、动物,或是海洋、陆地系统,优势生物都是少数特定、且全球分布的物种。举例而言,兽孔目的水龙兽,这种草食性动物的数量,占了当时陆地草食性动物约90%;以及双壳纲的克氏蛤( Claraia )、正海扇( Eumorphotis )、蚌形蛤( Unionites )、 Promylina 。正常的生态系统应由众多物种构成,并占据者不同的栖息地与生态位 。
在灭绝事件存活下来的某些劣势物种,数量与栖息地大幅增加。例如:双壳纲的海豆芽( Lingula )、出现于奥陶纪的叠层石、石松门的肋木( Pleuromeia )、种子蕨的二叉羊齿( Dicrodium )。
海洋生态系统的改变

在二叠纪末灭绝事件后,滤食性固着动物(例如海百合)的数量大幅减少
在二叠纪-三叠纪灭绝事件发生前,接近67%的海生动物是固着动物,附着于海床生存;在中生代期间,只有约50%的海生动物是固着动物,其余皆是可自由行动的动物。研究显示,事件发生后,栖息在海底表层的滤食性固着动物,例如腕足动物、海百合,物种数量减少;而构造较复杂的动物,例如蜗牛、海胆、螃蟹,物种数量增加。
在二叠纪末灭绝事件发生前,同时存在复杂与单纯的海洋生态系统,比例相当;生态系统经过复原后,复杂的海洋生物群落在数量上超过单纯的海洋生物群落,比例相当于3/1 ;这使海生掠食动物面临食物竞争的压力,促成中生代海洋生物的进一步演化。
在灭绝事件发生前,双壳纲是非常罕见的物种。在三叠纪,双壳纲成为数量众多、多样化的生物群;其中的厚壳蛤(Rudists),是形成中生代时期礁岩的主要生物。部分研究人员认为双壳纲的兴起,发生于两次灭绝高峰的间期 。
在灭绝事件中,海百合纲的物种数量、多样性大幅减少,但没有完全灭亡 。在灭绝事件后,海百合纲接着发生适应辐射,除了数量、栖息地增加,也发展出柔软的腕足、具可动性 。
陆地脊椎动物

在三叠纪早期,水龙兽是陆地上最繁盛的脊椎动物。
水龙兽属于兽孔目二齿兽类,是种体型接近猪的草食性动物,在三叠纪最早几个时期,占据了90%的陆地动物生态位 。除此之外,还有体型较小的肉食性犬齿兽类幸存,犬齿兽类是哺乳动物的祖先。根据非洲南部卡鲁盆地的化石纪录,有少数二叠纪物种也存活过二叠纪-三叠纪灭绝事件,例如兽头亚目的四犬齿兽( Tetracynodon )、麝喙兽( Moschorhinus )、 Ictidosuchoides ,但数量没有水龙兽繁盛 。
在三叠纪的早期,主龙类的数量远少于兽孔目;但在三叠纪中期,主龙类取代兽孔目的优势生态位 。在三叠纪中到晚期,主龙类的恐龙演化出现,并逐渐成为中生代的优势陆地动物 。兽孔目与主龙类的兴衰,与哺乳类的演化过程有关。兽孔目与随后出现的哺乳形类,在这个时期多半演化成小型、夜行性的食虫动物。夜间的生活习性,至少使哺乳形类发展出毛发与较高的代谢率 。
两栖动物离片椎目的某些物种,在灭绝事件中几乎灭亡,但快速复原。在三叠纪的大部分时间,虾蟆螈( Mastodonsaurus )与长吻迷齿螈下目(Trematosauria)是主要的水生与半水生掠食动物,以其他四足类、鱼类为食 。
陆地脊椎动物经过相当长的时间,才从二叠纪末灭绝事件复原。一位研究人员估计陆地脊椎动物花了3000万年,直到三叠纪晚期,才恢复之前的繁盛与多样性。此时的陆地脊椎动物包含:两栖动物、原始主龙类、恐龙、翼龙目、早期鳄鱼、以及哺乳形类 。
可能的灭绝原因
关于二叠纪-三叠纪灭绝事件的发生过程,目前已有多种假设,包含剧烈与缓慢的过程;白垩纪-第三纪灭绝事件的发生过程,也有类似的假设。剧烈过程理论的成因,包含大型或多颗陨石造成的撞击事件、连续性火山爆发、或是海床急骤释放出大量甲烷水合物。缓慢过程理论的成因,包含海平面改变、缺氧、以及逐渐增加的干旱气候 。
陨石撞击事件
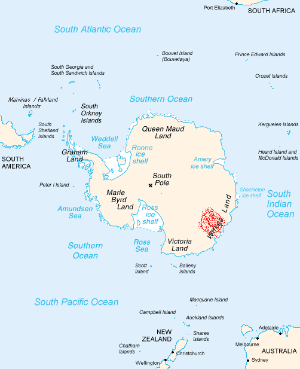
位于南极洲的威尔克斯地陨石坑,直径达500公里,形成时间在5亿年内,规模与时代使它成为本次灭绝事件的可疑成因,但证据不够明确。
白垩纪-第三纪灭绝事件由撞击事件造成的证据,促使科学家们推论其他灭绝事件由撞击事件造的的可能性,尤其是二叠纪-三叠纪灭绝事件。因此,科学家们尽力寻找那个时代的大型陨石坑与撞击证据。
目前已在部分二叠纪/三叠纪交界的地层,发现撞击事件的证据,例如:在澳洲与南极洲发现罕见的冲击石英 、富勒烯包覆的外太空惰性气体 、南极洲发现的玻璃陨石 ,以及地层中常见铁、镍、硅微粒 。但是,上述证据的真实性多受到怀疑 。在南极洲石墨峰(Graphite Peak)发现的冲击石英,经过光学显微镜与穿透式电子显微镜重新检验后,发现其中的结构并非由撞击产生,而是形变与地壳活动(例如火山)造成的 。
目前已经发现数个可能与二叠纪末灭绝事件有关的陨石坑,包含:澳洲西北外海的贝德奥高地 、南极洲东部的威尔克斯地陨石坑 。但没有可信服的证据,可证明这两个地形是由撞击产生。以威尔克斯地陨石坑为例,这个位在冰原下的凹地,年代无法确定,可能晚于二叠纪末灭绝事件才形成。其中最争议的陨石坑,是由地质学家Michael Stanton在2002年提出的,他主张墨西哥湾是形成于二叠纪末期的一次撞击事件,而该撞击事件也造成了二叠纪-三叠纪灭绝事件 。
如果二叠纪末灭绝事件的主因是由于撞击,很有可能陨石坑已经消失于地表。地球表面有70%是海洋,所以陨石或彗星撞击海洋的概率,是撞击陆地的两倍以上。但是,地球的海洋地壳会因聚合与隐没作用而消失于地表,所以目前无法找到距今2亿年以上的海洋地壳。如果当时有非常大型的撞击事件,撞击会使该处地壳破裂、变薄,造成大量的熔岩 。
撞击理论最受到关注的原因是,它可与其他灭绝现象产生因果连结 ,例如西伯利亚暗色岩火山爆发,可能由大型撞击产生 ,甚至是大型撞击的对跖点 。即使撞击发生于海洋,陨石坑因为隐没带而消失于地表,应会留下其他证据。如同白垩纪-第三纪灭绝事件,如果有撞击事件发生,会产生大量撞击抛出物,相同时代的许多地层会发现大量亲铁元素,例如铱。若灭绝事件由陨石撞击引发,可以解释在灭绝后,生物没有快速的适应演化。
火山爆发
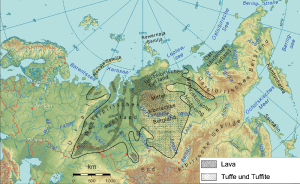
西伯利亚暗色岩的熔岩范围
在二叠纪的最后一期,发生两个大规模火山爆发:西伯利亚暗色岩、峨嵋山暗色岩。峨嵋山暗色岩位于现今中国四川省,规模较小,形成时间是瓜达鹿白阶末期,形成时的位置接近赤道 。西伯利亚暗色岩火山爆发是地质史上已知最大规模的火山爆发之一,熔岩面积超过200万平方公里。西伯利亚暗色岩火山爆发原本被认为持续约数百万年,但近年的研究认为西伯利亚暗色岩形成于2亿5120万年前(误差值为30万年),接近二叠纪末期 。
峨嵋山暗色岩与西伯利亚暗色岩火山爆发,可能制造大量灰尘与酸性微粒,遮蔽照射到地表的阳光,妨碍陆地与海洋透光带的生物进行光合作用,进而遭成食物链的崩溃。大气层中的酸性微粒,最后形成酸雨降落到地表。酸雨对陆地植物、可制造碳酸钙硬壳的软体动物与浮游生物造成伤害。火山爆发也释放大量二氧化碳,形成温室效应。大气层中的灰尘与酸性物质降落到地表之后,过量的二氧化碳持续形成温室效应 。
与其他火山相比,西伯利亚暗色岩更为危险。洪流玄武岩会产生大量的流动性熔岩,只会喷发少量的蒸气、碎屑进入大气层。但是,西伯利亚暗色岩火山爆发的喷出物质,似乎有20%是火山碎屑,这些火山灰与火山砾进入大气层后,会造成短时期的气候寒冷 。带有玄武岩的熔岩侵入碳酸盐岩、或带有大型煤层的地区,会产生大量的二氧化碳,会在大气层的灰尘降落到地表后,造成全球暖化 。
这些火山爆发事件的规模是否足以造成二叠纪末灭绝事件,仍有争议。峨嵋山暗色岩接近赤道区,火山爆发所制造的灰尘与酸性物质,会对全世界造成影响。西伯利亚暗色岩的规模较大,但位置在北极区内,或在北极区附近。如果西伯利亚暗色岩火山爆发的持续时间在20万年内,会使大气层中的二氧化碳达到正常程度的两倍。近年的气候模型显示,大气层中的二氧化碳含量加倍,会使全球气候上升1.5°C到4.5°C,这会造成严重的影响,但没有到二叠纪-三叠纪灭绝事件的严重程度 。
在2005年,日本NHK与加拿大国家电影委员会(NFB)制作的《地球大进化》( Miracle Planet ; 地球大進化〜46億年・人類への旅 )电视节目,提出火山爆发形成的轻微全球暖化,导致甲烷水合物的气化;由于甲烷气体对全球暖化的影响,是二氧化碳的45倍,甲烷水合物的气化进而导致不断循环的全球暖化。
甲烷水合物的气化
科学家们已在全球许多地点的二叠纪末碳酸盐矿层中,发现当时的碳13/碳12比例有迅速减少的迹象,减少了约10‰左右 。从二叠纪末开始,碳13/碳12比例发生了一系列的上升与下降现象,直到三叠纪中期才稳定、停止;而发生于二叠纪末的第一次变动,是其中规模最大、最迅速的变动。在三叠纪中期,碳13/碳12比例稳定之后,可制造碳酸钙外壳的动物开始复原 。
碳13/碳12比例的下降,可能有以下多种因素 :
火山爆发产生的气体,其碳13/碳12比例低于正常值约5~8‰。但若要使全球的碳13/碳12比例下降约10‰,其火山爆发的规模将超越目前已知的任何地质纪录 。
生物活动降低,使环境中的碳12更慢被摄取,更多碳12进入沉积层,而使碳13/碳12比例降低。所有的化学反应是建立在原子间的电磁力。较轻的同位素,其化学反应较快。所以生物化学过程会用到较轻的同位素。但关于古新世-始新世交替时期最大热量(PETM)事件的研究发现,该时期的碳13/碳12比例小幅降低约3到4‰。根据假设,即使将全部的有机碳(包含生物、土壤、海洋)进入土壤沉积层中,也不会达到古新世/始新世交替时的小幅度碳13/碳12比例下降。由此可知生物活动降低不是二叠纪末期的碳13/碳12比例降低的原因 。
埋在沉积层中的死亡生物,体内的碳13/碳12比例小于正常值约20到25‰。就理论上而言,如果海平面迅速降低,浅海地区的沉积层曝露到空气后,开始氧化作用。但若要使全球的碳13/碳12比例下降约10‰,要有6.5到8.4兆吨有机碳经氧化后形成,而沉积层本身需要数十万年的氧化。这似乎不太可能发生 。
间歇性海洋高氧与缺氧事件,也可能是三叠纪早期的碳13/碳12比例下降的原因 。全球性的海洋缺氧现象,本身也是灭绝事件的原因之一。二叠纪末期到三叠纪早期的陆地,多为热带地区。热带的大型河流会将沉积层中有有机碳带入海洋,尤其是低纬度的邻近海盆。生物化学过程会用到较轻的同位素,所以有机碳的碳13/碳12比例低。大量有机碳的迅速(以相对而言)释放与沉降,可能会引发间歇性的高氧/缺氧事件 。元古宙晚期到寒武纪交接时期,也曾发生碳13/碳12比例的下降,可能与此相关,或是其他与海洋相关的因素 。

甲烷水合物即俗称的可燃冰,大部分位在大陆棚,被视为未来的潜在能源之一。
其他的理论则有:海洋大量释放二氧化碳 、以及全球的碳循环系统经历长时间的重整 。
但最有可能导致全球性碳13/碳12比例下降的因素,是甲烷水合物气化产生的甲烷 。而利用碳循环模型模拟的结果,甲烷最有可能导致如此大幅的下降 。甲烷水合物是固态形式的水于晶格(水合物)中包含大量的甲烷。甲烷是由 甲烷菌 ( 英语 : Methanogen ) 制造,碳13/碳12比例低于正常值约60‰。甲烷在特定的压力与温度下,会形成包合物,例如永冻层的近表层,并在大陆棚、更深的海床等地区大量形成。甲烷水合物通常出现在海平面300米以下的沉积层。最深可在水深2000米处发现,但大部分在水深1100米以上 。
西伯利亚暗色岩火山爆发产生的熔岩面积,是以往认定的两倍以上,新发现的熔岩地区,在二叠纪末时几乎是浅海。极有可能这些浅海地区蕴含甲烷水合物,而火山爆发产生的熔岩流入海床后,促使甲烷水合物的汽化 。
由于甲烷本身是种非常强的温室气体,大量的甲烷被视为造成全球暖化的主要原因。证据显示该时期的全球气温上升,赤道区上升约6°C,高纬度地区上升更多。例如:氧18/氧16比例的下降 、舌羊齿植物群(舌羊齿与生存于相同地区的植物)消失,由生存于低纬度的植物群取代 。
碳13/碳12比例的变动,被认为与大量释放的甲烷有关,但两者在三叠纪早期的变动模式,并不吻合。要造成如此大的气候变迁,所需要的甲烷量,是引发古新世-始新世交替时期最大热量的五倍 。但三叠纪早期曾出现大量碳13,使碳13/碳12比例迅速增加,而后下降 。
海平面改变
当原本沉浸的海床露出海平面时,会造成海退。海平面的下降会使浅海的生存区域减少,破坏当地的生态系统。浅海的可栖息地,富含食物炼下层的生物,这些生物的减少,使赖其以维生的生物竞争食物更激烈 。海退与灭绝事件之间似乎有部分关联,但另有证据认为两者间没有关系,而海退会形成新的栖息地。 海平面的变化,同时也导致海底沉积物的变化,并影响海水温度与盐度,进一步造成海生生物的多样性衰退 。
海洋缺氧
有证据显示,二叠纪末期的海洋发生了缺氧事件。在格陵兰东部的一个二叠纪末期海相沉积层,指出当时有明显、快速的海洋缺氧现象 。而数个二叠纪末沉积层的铀/钍比例,也指出在这次灭绝事件发生时,海洋有严重的缺氧现象 。
缺氧事件可能导致海洋生物的大量死亡,只有栖息于海底泥层、可以进行无氧呼吸的细菌不受影响。另有证据显示,这次海洋缺氧事件,造成海床大量释放硫化氢。
海洋缺氧事件的原因,可能是长时间的全球暖化,降低赤道区与极区之间的温度梯度,进而造成温盐环流系统的缓慢,甚至停止。温盐环流系统的缓慢或停止,可能使得海洋中的含氧量减少 。
但是,某些研究人员架构出二叠纪末期的海洋温盐环流系统,认为当时的温盐环流系统无法解释深海区域的缺氧现象 。
硫化氢
二叠纪末期发生的海洋缺氧事件,可能使 硫酸盐还原菌 ( 英语 : Sulfate-reducing bacteria ) 成为海洋生态系统中的优势物种,包含脱硫杆菌目、脱硫弧菌目、互营杆菌目、热脱硫杆菌门,这些生物会制造大量的硫化氢,过量的硫化氢会对陆地、海洋中的动植物造成毒害,并破坏臭氧层,使生物暴露在紫外线下 。在二叠纪末到三叠纪早期发现许多绿菌,它们进行不产氧光合作用,释放出硫化氢。绿菌的兴盛时期,与二叠纪末灭绝事件和事后的长期复原,时期相符 。大气层中的二氧化碳增加,植物却大规模灭亡,硫化氢理论可以解释植物的大规模灭亡。二叠纪末地层中的孢子化石,多数带有不正常特征,可能是由硫化氢破坏臭氧层,大量的紫外线进入地表造成。
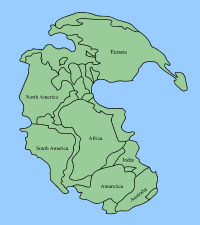
盘古大陆的地图
盘古大陆的形成
在二叠纪中期(约是乌拉尔世的空谷阶),几乎地表的所有大陆聚合成盘古大陆,盘古大陆由泛大洋环绕着,而东亚部分直到二叠纪末期才与盘古大陆聚合 。盘古大陆的形成,使得全球大部分的浅水区域消失,而浅水区域是海洋中最多生物栖息的部分。原本隔离的大陆架连接之后,使彼此独立的生态系统开始互相竞争。盘古大陆的形成,造成了单一的海洋循环系统,以及单一的大气气候系统,在盘古大陆的海岸形成季风气候,而广大的内陆则形成干旱的气候。
在盘古大陆形成后,海洋生物数量减少,灭亡比例接近其他大型灭绝事件。盘古大陆的形成,似乎对陆地生物没有造成严重的变化,二叠纪晚期的兽孔目反而因此扩大生存领域、更为多样性。盘古大陆的形成,使海洋生物开始减少,但不是导致二叠纪末灭绝事件的直接原因。
多重原因
二叠纪-三叠纪末灭绝事件的产生原因,可能由上述事件连锁、交错形成,并日趋严重。西伯利亚暗色岩的火山爆发,除了产生大量的二氧化碳与甲烷,也破坏邻近地区的煤层与大陆架 。接下来的全球暖化,间接导致地质历史中最严重的海洋缺氧事件。海洋的缺氧,使硫化菌等进行不产氧的化合作用生物兴起,它们释放出大量的硫化氢 。
但是,这连锁、交错的事件,部分环节相当薄弱。碳13/碳12比例的变动,被认为与大量释放的甲烷有关,但两者在三叠纪早期的变动模式,并不吻合 。二叠纪末期的海洋温盐环流系统,不会造成深海区域的缺氧事件 。
延伸阅读
(英文) Over, Jess (editor), Understanding Late Devonian and Permian–Triassic Biotic and Climatic Events , (Volume 20 in series Developments in Palaeontology and Stratigraphy (2006). The state of the inquiry into the extinction events.
(英文) Sweet, Walter C. (editor), Permo–Triassic Events in the Eastern Tethys : Stratigraphy Classification and Relations with the Western Tethys (in series World and Regional Geology)
科学网:2.5亿年前两大物种灭绝之谜获突破性进展
BBC:二叠纪末灭绝事件的复杂原因
BBC2:在2.5亿年前,地球上的生物几乎灭亡
SpaceRef:南极洲冰层下的陨石坑(影像由俄亥俄州立大学提供)
Science Daily:全球暖化使大气层充满硫化氢,并导致二叠纪末灭绝事件
Science Daily:南极洲的冰层下发现疑似陨石坑的大坑洞
SPACE.com:南极洲发现地表最大的陨石坑,可能是地质年代中最严重的生物灭绝有关]
Space.com:地层中的二叠纪末生物灭绝证据
NAI:二叠纪灭绝事件是否由撞击事件导致?
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。





- 有价值
- 一般般
- 没价值
















推荐阅读





关于我们

APP下载


