






支序分类学
术语
一个特症状态若是同时表现在外类群和其祖先身上时,称之为“祖征”;而若只出现在之后的后代时,则称之为“衍征”。用祖征和衍征来代替原始和先进这些词是因为,若是在不同的环境之下,每种特征都有可能是先进的。用祖征来指分支的“分类基础”在非正式的情况下并不会不普遍。
一些种类如果比其他种类具有更多的祖征,称为基本分支,一般来说基本分支比其他衍生分支的种类要少得多,基本分支也可能现在已经不存在,例如鸟类和哺乳动物的共同祖先就已经不存在,虽然它们之间有许多共同的祖征存在。
支序分类方法
分支分类分析必须收集信息,并区分什么是“一般特征”什么是“特殊性状”。在以前,研究者必须区分哪些特殊性状是共同祖先以前具有的(祖征),哪些是共同祖先单独具有的特征(共源性状),要和分支以外的种群比较,只有共源性状才能用来确定一个分支。
要确定一个分支,必须有足够大量的共源性状,才能说明这种特征不是由于共同的祖征,不是由于趋同进化造成的同形相似(因为生活在同一环境,而不是因为有同一祖先造成的相似特征)。例如鸟类和昆虫有生活在相似的环境中都具有的翅膀(共同的特征),但却是沿着不同的进化路线发展的,如果将它们放到一个分支内就是错误的。
共源特征确定后,要依照简约化的原理确定之间的关系(也是一种奥卡姆剃刀原理),另外也主要也依照分子进化理论,选取最接近的分支。
现代不太容易出现由于研究者自身造成的偏差,因为一般分析都使用计算机软件,现在有各种复杂的软件可以用来分析和计算分支。
但有时也会出现许多冲突,例如外形特征和基因分析完全不同等,造成分到不同的分支内,现在种系发生学提供了得到大部分人赞同的结论。
现代DNA系列的测定已经比较容易,种系发生学得到分子数据的支持,计算机系统可以处理大量的数据,能精确筛选出真正的共源特征数据。更为有效的方法是利用染色体中的反转座子记号。最理想的是将形态学、分子生物学、以及行为生态等种系发生学的信息结合起来,没有一种方法是完全完美的。所有的方法都有自己的误差,在形态学中经常发生趋同进化的错误,但在分子生物学中却经常有和外在特征冲突的现象,分子数据虽然比利用化石数据更精确,但也充满误差。各种方法的比较和结合,才能得到更好的结果。
亲缘分支理论并不能取代现在所有的进化理论,只是对现有的知识进行一些修正。这种理论也可以使用到生物学以外的领域,目前已经应用到历史语言学中确定语言的族属分类,和古代文献考订中确定文献派生情况等。
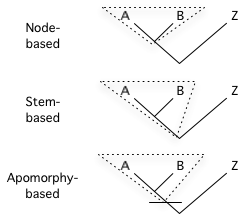
亲缘分支法所用的三种分支方法 结点分支 :最近的共同祖先分出的A和B以及它们的后代。 主干分支 : 包括A和B 的比较远的共同祖先及其后代,但不包括Z。 派生分支 : A 和B 最近的共同祖先及其后代,还包括具有一定的派生特性的较远祖先。
支序分类法分类
自从1960年代以来,生物界出现倾向用亲缘分支分类法的“分支主义”,他们主张取消所有非分支的分类系统。但其他分类学家认为虽然种系发生学要应用亲缘分支的技术,但分类仍然有使用传统的门类的必要,两种分类法都应该存在,因为不仅存在单源种系,也有同源种系。
单源种系可以应用分支,全体种系都是从一个祖先派生出来的。同源种系虽然意义相同,但其中一部分种群有独立的进化道路,产生很大的区别。例如鸟类和爬行类具有同一祖先,但进化道路相当不同。脊椎动物和无脊椎动物也是从共同祖先演化来的,但不能分到同一类中。
此外还有多源种系,例如以前分到一类“厚皮类”中的动物象和犀牛并不是来源于同一祖先,而是各自由不同的薄皮祖先进化来的。演化分类学者认为多源在分类学中是错误的,如同将趋同演化的动物或同形的动物分到一类中一样。
分支学者认为同源种系和多源种系是一样错误的,单系群是依据共源性状,可以将各种群进行客观地分类。而同源和多源是依据外观特征,容易造成主观的错误。但其他学者认为分支法将种群分类从“低级”向“高级”分类,容易导致目的论,目的论是在演化研究中应该避免的。
分支学者认为传统的分类方法人为地将物种分成不同的层次,不能真正代表物种演化的实际情况,因此应该放弃林奈的分类法,全部使用简单的亲缘分支层次。
而其他有不同意见的学者认为所有的分类方法都是人为的,都有主观因素在内,所有物种都是在持续不断地演化,任何分类划分都是人为制造前半部单一起源,后半部同源发展,在树形结构的早期就应用同源分类是必要的。例如发展成为灵长类的早期脊椎动物不能放到任何单源分支中。他们还认为同源分类提供了生物形态、生态即生命演化历史中重要变化的信息。也就是说传统分类和分支分类都很有价值,但在不同方面是为不同目的而起作用。
相关条目
系统发生树
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。


- 有价值
- 一般般
- 没价值
















推荐阅读

关于我们

APP下载


