






始新世
下分
始新世经常被分为早期和晚期,亦经常被分为早期、中期和晚期。其对应的岩层分别为下始新世岩层、中始新世岩层和上始新世岩层。根据不同的动物的活跃,始新世又可以细分为:
下始新世岩层主要由伊普雷斯阶构成,有时还包括卢台特阶;上始新世岩层主要由普里阿邦阶构成,有时还包括巴尔顿阶;此外,卢台特阶和巴尔顿阶则构成中始新世岩层。
气候
由于被称为 PETM ( 英语 : 古新世-始新世极热事件 ) ,是地球地质历史记录现的最快的、强度最大的一次全球变暖事件,地球温度迅速升高,始新世就此开始。这次迅速和剧烈的升温(在高纬度地区温度上升了7°C 之多)持续的时间少于10万年 ,但是导致了大量物种急剧灭绝,从而形成了古新世和始新世生态系统的分野。
始新世的全球气候可能是新生代中区域变化幅度最小的;从赤道到极地的气温变化幅度只有今天的一半,而深海洋流则异常温暖。 极地地区比现在温暖得多,接近于现今西北太平洋的温度;温带森林已经扩展到了极地地区,同时多雨的热带气候区则延伸至北纬45°地区。温带地区的气候与现在相比大有不同,而热带地区的气候则可能接近于今世。 不过最近在哥伦比亚发现的一种可能生活于始新世的巨蛇(约有校车大小)表明,可能相反的,当时的热带比现今温暖的多 ,这也与计算机模拟出的始新世气候相符 。
尽管始新世的全球气候仍然比较温暖,但也正是在始新世后期,开始了全球变冷的缓慢趋势,这可能是由发生于北冰洋的绿萍事件以及在冈瓦纳古陆最终破碎之后形成的南极洲环流共同造成的。这种变冷趋势最终导致了更新世冰期的出现。
古地理
File:Paleogene-EoceneGlobal.jpg 约5,000万年前,始新世时期的全球古地理复原图
在始新世,各大洲继续飘移向它们目前所在的位置。
始新世初始,澳大利亚和南极洲仍然相连,同时温暖的赤道洋流汇入寒冷的南极水域,使得热量在全球范围内获得分配,从而保持全球的较高温度。但是在4500万年前,当澳大利亚从南方大陆中分裂出来时,温暖的赤道洋流开始偏离南极地区,在两块大陆之间形成了一个孤立的寒冷水道。南极地区持续变冷,南极水域开始结冻,并向北方输送冷水和海冰,使寒冷局势进一步加剧。
北方的超级大陆——劳亚古大陆也开始分裂,欧洲、格陵兰岛和北美洲相继从中分裂而出。
在北美洲西部,于始新世开始了造山运动,在平原的抬升过程中形成了诸多巨大的湖泊,并进而沉积形成了绿河岩层。
在欧洲,阿尔卑斯山脉的抬升将特提斯洋的最后残迹——地中海包围,从而使特提斯洋最终消失,并形成了另外一个浅海,这个浅海的北部区域分布着一系列的群岛。虽然北大西洋正在扩张,但是北美洲和欧洲之间可能仍有陆桥相连,因为两块大陆上的动物区系十分相似。
印度次大陆继续漂离非洲大陆,并开始撞击亚洲大陆,喜马拉雅山脉开始形成。
假说认为始新世温暖的全球气候是由深埋于大洋底部的甲烷气水包合物被大量释放引起的失控的温室效应造成的。该种包合物本埋于泥层之下,但是受到了大洋升温的扰动。甲烷的全球变暖潜能(即暖化能力)比二氧化碳高22倍。
动物

中爪兽,一种肉食性的有蹄类哺乳动物
大量的现代哺乳动物化石最早出现于始新世早期。在始新世开始之初,一些新哺乳动物群到达北美。这些原始的哺乳动物,如偶蹄类动物,奇蹄类动物和灵长类动物,特征是细长的四肢,有抓取物体的能力,可以透过鉴别牙齿来区分它们的食性。小型化是主要发展趋势,始新世所有新产生的哺乳类动物都是小型的,重量轻于10千克;且基于牙齿尺寸的关系,始新世的哺乳动物的体型只有原始的古新世哺乳动物的百分之六十。它们的体型也比其后的哺乳类动物小。人们认为始新世时期较高的温度比较适宜小型哺乳类动物的生存,因为较之大型哺乳类动物,小型哺乳类动物更容易解决散热的问题。
古新世时期的古老灵长类动物于始新世衰落了,而其生态环境中的许多适当的场所都被更有效的啮齿类动物所占据。始新世中期的脊椎动物群不像始新世早期的那般分布在世界各地;因此所形成的隔离状态,使不同的进化趋势得以出现在北美和欧洲的有蹄类动物群中。许多现代哺乳类动物的始祖都已经出现,包括蝙蝠、长鼻目动物、灵长目动物、啮齿类动物和有袋类动物。在北美洲西部、欧洲、巴塔哥尼亚、埃及和东南亚都发现了重要的始新世时期的陆生动物化石;海洋动物化石则在南亚和美国东南部有大量发现。爬行动物,如蟒蛇和海龟的化石也十分丰富。近年来在哥伦比亚发现了如校车般大小的巨蛇残骸 ,这种巨蛇在如现今这般温暖的赤道是无法生存下去的,印证了之前依据其他证据提出的高温观点 ("a snake of this size would require a minimum mean annual temperature of 30–34 °C to survive")。
在始新世,植物和海洋动物区系都和现今十分接近。许多现代的鸟类在始新世首次出现。一些科学家相信的第一种灵长目动物出现在5,500万年前伊普雷斯阶时期;但是分子钟和新的古生物学发现都表明第一种灵长目动物可能出现得更早,大约在9,000万年前的白垩纪。

呈微球型和显球型的有孔虫门生物化石;生活在始新世时期的阿拉伯联合酋长国地区;毫米级别
海洋动物
始新世的海洋温暖而充满鱼群和其他海洋生物。 出现了:
最早的真鲨目动物。
早期的海生哺乳动物:包括龙王鲸(早期的一种鲸鱼,被认为由一种生活于始新世早期的陆生动物进化而来)、最早的海牛目动物。
植物
在始新世早期,因为高温的气候以及温暖海洋,所以创造了一个潮湿而温和环境。除了干旱的沙漠之外,地表完全被森林所覆盖。
极地地区的森林面积广大。在北极地区的埃尔斯米尔岛上发现了始新世时期的树木化石以及保存至今的树木,如落羽杉和水杉。这些保存至今的树木不是化石,而是原先存在于沼泽森林缺氧的水中,并在未及腐烂的情况下被掩埋的实物。即使在那时,埃尔斯米尔岛也只比今天所在的位置低了几个纬度。另外,在格陵兰岛和阿拉斯加都发现了始新世时期的温带森林,甚至是热带植被的化石。热带雨林则扩展至太平洋西北岸和欧洲地区。
在始新世早期,棕榈树分布范围北及阿拉斯加和北欧地区,不过其后气候变冷,棕榈树在这些地区渐趋衰落。水杉在这一时期也分布广泛。
在始新世中期开始的气候变冷,致使到了始新世末期时大陆内部开始变得干燥,在某些地区,森林分布区域萎缩。此时新发展起来的草原也仅限于河岸和湖畔地区,还未扩展至平原地区和现今的热带或带大草原地区。
气候变冷亦导致了季节变化的出现。落叶树种能够更好的适应剧烈的温度变化,于是比之常绿树种,开始占有优势。在始新世末期,落叶林覆盖了北方大陆的大片区域,包括北美洲、欧亚大陆和北极地区;雨林则只分布在了赤道和近赤道的南美洲、非洲、印度次大陆和澳大利亚。
在始新世开始之时,南极大陆上还覆盖着大片的温带雨林,乃至是带雨林,但是随着时间流逝,气候变得越来越寒冷;喜热的热带植被消失了,到渐新世开始之时,这块大陆上的植被变成了落叶林和广袤的苔原。
大置换
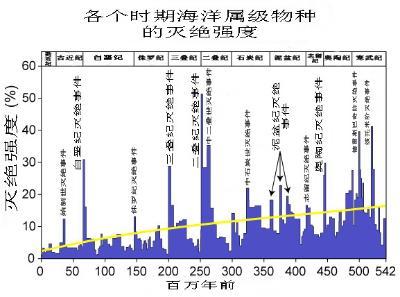
始新世灭绝事件,纵轴为灭绝的科数占地球的百分比。
大置换,即物种连续性上的“大断裂” ,是3,350万年前欧洲范围内的一次物种大灭绝,标志着始新世的最后一个阶段——普里阿邦阶的结束,此后,亚洲的物种大量迁入欧洲。大置换以大范围的物种灭绝和小规模、孤立孑遗种群的异地物种形成为标志 ;这个名称由瑞士古生物学家汉斯·乔治·斯特林(Hans Georg Stehlin)所取 ,被用以形容始新世和渐新世交迭期间欧洲地区哺乳类动物的大规模灭绝。而在亚洲发生的类似规模的物种灭绝则被称为“蒙古重建”(Mongolian Remodelling)。
大置换标志着在亚洲物种大规模迁入之前,欧洲区域性动物区系进化链的断裂。J·J·胡克(J.J.Hooker)及其团队如此概括这次灾难:
大置换前欧洲的动物区系主要由古兽科动物(马的远亲)、偶蹄类动物的6个科(无防兽科、剑齿兽科、河猪科、长尾猪科、双锥齿兽科和疑刍驼科)、啮齿类动物Pseudosciuridae,以及灵长类动物Omomyidae和Adapidae组成。
大置换后欧洲的动物区系中包括了一种最早的犀牛(属犀科)、3个偶蹄动物科(巨猪科、炭兽科和吉洛鹿科,分别于现今的猪、河马和反刍类动物有亲缘关系)、多种啮齿科动物(如Eomyidae、仓鼠科、河狸科)以及无盲肠动物中的刺猬科动物。而原来的古兽马(Palaeotherium)、无防兽科动物、剑齿兽科动物和疑刍驼科动物则完全消失了。
只有有袋类中的疱负鼠科(Herpetotheriidae)动物、偶蹄类中的新兽科动物以及啮齿类中的Theridomyidae科动物和睡鼠科动物熬过了这次灾难,种群并未衰退。
许多科学家认为大置换并非是由单一的原因造成的,不管这个原因是与初期的极地冰蚀 和海平面下降相联系的气候变化,还是来自亚洲的物种的竞争。另一个假说是一颗或多颗撞击向西伯利亚和切萨皮克湾的火流星造成了这次物种大灭绝。与这一事件相关的、发生在之前的一系列全球件则进一步证实大置换发生于渐新世最早期,大约比亚洲首批迁徙物种进入欧洲早35万年。
大置换的变化模型中的一个重要支撑要素是——在大置换中,欧洲地区所有的灵长类动物都灭绝了;但是最近发现的一种生活于渐新世早期的、大小与鼠类相仿的灵长类动物——海岸潟湖灵长类(omomyid) (这一发现表明在那个时期,小型哺乳类动物更易于生存),则进一步削弱了这一大置换变化模型。
参见
地质时代表
显生宙
新生代
第三纪
古近纪
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

相关资料
展开



- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读
知识互答

关于我们

APP下载


