






ATP酶
功能
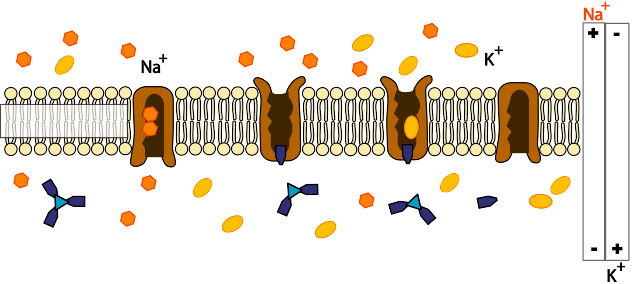
钠钾ATP酶的功能图示
跨膜ATP酶可以为细胞输入许多新陈代谢所需的物质并输出毒物、代谢废物以及其他可能阻碍细胞进程的物质。例如,钠钾ATP酶(又称为钠/钾离子ATP酶)能够调节细胞内钠/钾离子的浓度,从而保持细胞的静息电位;氢钾ATP酶(又称为氢/钾离子ATP酶或胃质子泵)可以使胃内保持酸化环境。
除了作为离子交换器,跨膜ATP酶还有其他类别,包括共转运蛋白(co-transporter)和“泵”(也有部分“离子交换器”也被称为“泵”)。这些跨膜ATP酶中,有一些可以造成膜内外电荷的流动,其他的则不行,因此又可以将这些转运蛋白分为生电型(electrogenic)和非生电型。
反应机制
与ATP水解反应耦合的转运是一个严格的化学反应,即每分子ATP水解能够使一定数量的溶液分子被转运。例如,对于钠钾ATP酶,每分子ATP水解能够使3个钠离子被运出细胞,同时2个钾离子被运入。
跨膜ATP酶需要ATP水解所产生的能量,因为这些酶需要做功:它们逆着热力学上更容易发生的方向来进行物质运输,换句话说,以膜为参照,它们可以将物质从低浓度的一边运送到高浓度的一边。这一过程被称为主动运输。
ATP合酶
三磷酸腺苷合酶是一类线粒体与叶绿体中的合成酶,它可以利用膜内外质子的浓度差作为能量源,将ADP与磷酸合成为ATP。ATP合酶这一作用正好与ATP酶相反。
同时,ATP合酶也可以催化逆反应,即ATP的水解。因此,从某种意义上来说,ATP合酶也是一类ATP酶。
相关人类基因
钠/钾离子转运: ATP1A1 , ATP1A2 , ATP1A3 , ATP1A4 , ATP1B1 , ATP1B2 , ATP1B3 , ATP1B4
钙离子转运: ATP2A1 , ATP2A2 , ATP2A3 , ATP2B1 , ATP2B2 , ATP2B3 , ATP2B4 , ATP2C1
镁离子转运: ATP3
氢/钾离子转运: ATP4A , ATP4B
线粒体中的氢离子转运: ATP5A1 , ATP5B , ATP5C1 , ATP5C2 , ATP5D , ATP5E , ATP5F1 , ATP5G1 , ATP5G2 , ATP5G3 , ATP5H , ATP5I , ATP5J , ATP5J2 , ATP5L , ATP5L2 , ATP5O , ATP5S
溶酶体中的氢离子转运: ATP6AP1 , ATP6AP2 , ATP6V1A , ATP6V1B1 , ATP6V1B2 , ATP6V1C1 , ATP6V1C2 , ATP6V1D , ATP6V1E1 , ATP6V1E2 , ATP6V1F , ATP6V1G1 , ATP6V1G2 , ATP6V1G3 , ATP6V1H , ATP6V0A1 , ATP6V0A2 , ATP6V0A4 , ATP6V0B , ATP6V0C , ATP6V0D1 , ATP6V0D2 , ATP6V0E
铜离子转运: ATP7A , ATP7B
I型,第8类: ATP8A1 , ATP8B1 , ATP8B2 , ATP8B3 , ATP8B4
II型,第9类: ATP9A , ATP9B
V型,第10类: ATP10A , ATP10B , ATP10D
VI型,第11类: ATP11A , ATP11B , ATP11C
非胃中的氢/钾离子转运: ATP12A
第13类: ATP13A1 , ATP13A2 , ATP13A3 , ATP13A4 , ATP13A5
参见
三磷酸腺苷
AAA蛋白
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。


- 有价值
- 一般般
- 没价值
















推荐阅读


关于我们

APP下载


