






RNA聚合酶
控制转录
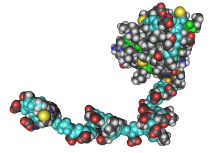
人类RNA聚合酶I、Ⅱ及Ⅲ的必要亚基
控制转录过程会影响基因表达的模式,并从而容许细胞适应不同的环境、执行生物内独特的角色及维持生存所需的代谢过程。所以,RNA聚合酶是活动不单是复杂,而且是有高度规律的。在大肠杆菌中,已确认超过100个因子可以修饰RNA聚合酶的活动。
RNA聚合酶可以在特定的DNA序列,称为启动子发动转录。它继而产生RNA链以补足DNA的模板股。并会加入核苷酸至RNA股,这个过程称为“延伸”。在真核生物的RNA聚合酶可以建立一条长达240万个核苷的链(等同于肌萎缩蛋白基因的总长度)。RNA聚合酶会优先在基因末端已编码的DNA序列(称为终结子)释放它的RNA转录本。
核糖体会把一些RNA分子会作为蛋白质生物合成的模板。其他会折叠成核酶或转运RNA(tRNA)分子。第三种可能性是RNA分子会单纯地用作控制调节将来的基因表达。(参考小干扰性RNA)
RNA聚合酶完成一个全新的合成。它能够这样造是因为它与起始的核苷酸独特的相互作用,能把它牢牢地抓住,方便对进入的核苷酸进行化学攻击。这种独特的相互作用解释了为何RNA聚合酶较喜欢以三磷酸腺苷(ATP)作为转录的开始,依次其次是三磷酸鸟苷(GTP)、三磷酸尿苷(UTP)及三磷酸胞苷(CTP)。与DNA聚合酶相反,RNA聚合酶包含了解旋酶的活动,所以无须另外的酶来卷开DNA。
细菌的RNA聚合酶
在细菌中,相同的酶催化三种RNA的合成:信使RNA(mRNA)、核糖体RNA(rRNA)及转运RNA(tRNA)。
RNA聚合酶是相对大的分子。核心酶有5个亚基(~400 kDa):
α 2 :这两个α亚基组合成酶及辨认调节因子。每个亚基有两个区,αC末端区及αN末端区,分别与启动子结合及与聚合酶的其他部分结合。
β:有着聚合酶的活动,负责催化RNA的合成。
β":与DNA结合。
ω:还未清楚它的功能。但是它在耻垢分枝杆菌中似乎是提供保护功能予β"亚基 。
为着与启动子的特定区域结合,核心酶须有其他亚基,称为σ。σ因子大大减低RNA聚合酶与非特定的DNA的关系,视乎σ因子本身而增加对某些启动子区域的独特性。所以完整的全酶有着6个亚基:α 2 、β、β"、σ及ω(~480 kDa)。RNA聚合酶的结构就有一个长约55Å(即5.5纳米)的沟道及直径为25Å(2.5纳米)。这个沟道正好适合20Å(2纳米)的DNA双股。55Å的长度可以接受16核苷酸。
当不使用时,RNA聚合酶会与弱结合部位结合,等待活性启动子的位点开启并快速转换。RNA聚合全酶所以在不使用时不是在细胞内自由浮动的。
真核生物的RNA聚合酶
真核生物有着几种RNA聚合酶:
RNA聚合酶I合成核糖体RNA(rRNA)前体45S,当成熟后会成为28S、18S及5.8S核糖体RNA,是将来核糖体的主要RNA部分。
RNA聚合酶Ⅱ合成信使RNA(mRNA)的前体及大部分小核RNA(snRNA)以及微型RNA(microRNA)。因为它在转录过程中需要多种转录因子才能与启动子结合,所以这是现时最多研究的种类。
RNA聚合酶Ⅲ合成转运RNA(tRNAs)、rRNA 5S及其他可以在细胞核及原生质找到的细小的RNA。
其他在线粒体及叶绿体的RNA聚合酶种类。
古菌的RNA聚合酶
古菌的RNA聚合酶只有一种,负责所有RNA的合成。古菌RNA聚合酶在结构和催化机理上与都与细菌、真核生物的聚合酶类似,尤其类似于真核生物的RNA聚合酶Ⅱ 。
古菌RNA聚合酶的研究开展较晚,第一项成果发表于1971年,the RNAP from the extreme halophile Halobacterium cutirubrum was isolated and purified. Crystal structures of RNAPs from Sulfolobus solfataricus and Sulfolobus shibatae set the total number of identified archaeal subunits at thirteen.
病毒的RNA聚合酶
很多病毒都有为RNA聚合酶编码。相信最多研究的病毒RNA聚合酶是噬菌体T7。它的RNA聚合酶是单一亚基的,与在线粒体及叶绿体所找到的RNA聚合酶相关,并且与DNA聚合酶同源。因此很多人相信大部分的病毒聚合酶是从DNA聚合酶演化而来,并不是直接与上述的多亚基聚合酶有所关联。
病毒聚合酶是繁杂的,且包括一些形态可以使用RNA(而非DNA)作为模板。反链核糖核酸病毒及双链核糖核酸病毒都是以双股RNA形式生存。但是,有些正链核糖核酸病毒,如小儿麻痹病毒,亦包含这些RNA依赖性RNA聚合酶。
转录辅助因子
有部分蛋白质可以与RNA聚合酶结合,并修饰其活动。例如大肠杆菌的greA及greB可以促进RNA聚合酶劈开接近链末端RNA模板的能力。这可以夺回陷入了的聚合酶分子,并且可以校对RNA聚合酶偶然的错误。另一种辅助因子Mfd涉及在转录合并修复中,而其他辅助因子则都是负责调节作用,即帮助RNA聚合酶选择是否表现某些基因。
纯化
RNA聚合酶可以用以下方式纯化:
磷酸纤维素柱层析
甘油梯度离心
DNA柱层析
离子交换柱
一并使用磷酸纤维素柱层析加上DNA柱层析
内部链接
DNA聚合酶
T7核糖核酸聚合酶
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。


- 有价值
- 一般般
- 没价值
















推荐阅读



关于我们

APP下载


