






爬行动物
爬行动物的分类

脊椎动物的简易演化树。传统分类上的爬行动物其实是并系群。如果包含鸟类、哺乳动物在内,则成为单系群羊膜动物,剔除哺乳类一支(合弓类)则相当于蜥形纲
分类历史
在1758年,瑞典生物学家卡尔·林奈建立二名法后几年,他根据部分蛇、蜥蜴生存于水中,而建立两栖纲,以包含两栖动物、爬行动物 。在1768年,自然学家Josephus Nicolaus Laurenti建立 爬行纲 (Reptilia),但当时两栖动物、爬行动物的界定仍然模糊 ; Repere 在拉丁语中意为“爬行” 。直到18世纪末期,两栖动物、爬行动物才被视为不同的动物。在1825年,拉特雷耶·皮埃尔·安德烈(Pierre André Latreille)建立“Batracia”纲,以取代两栖纲,并将四足动物分成四大类:两栖动物、爬行动物、鸟类、哺乳动物,已经相当接近现代传统分类法 。
在托马斯·亨利·赫胥黎的推广下,Latreille的分类逐渐普及。理查·欧文则将似哺乳爬行动物包含在爬行纲的范围。
在传统的分类法中,爬行动物的范围为羊膜动物除去鸟类与哺乳类。因此爬行纲被定义为包含鳄鱼、短吻鳄、喙头蜥、蜥蜴、蛇、蚓蜥、以及乌龟在内的动物群。这个名称与定义成为后来常见的用法。然而在最近几年,许多分类学家主张生物学分类应该为单系群,这些分类应该包含特定物种的所有后代。上述的爬行纲定义因为不包含哺乳纲与鸟纲,而被视为并系群,它们都是原始爬行动物的后代。
哺乳类是个演化支,因此亲缘分支分类法承认传统的哺乳纲分类;而鸟类也是个演化支,正式的名称为鸟纲。事实上,哺乳纲与鸟纲是羊膜动物的次演化支。但传统分类法中的爬行纲并不是一个演化支,它其实是部分的羊膜动物,不包含哺乳纲与鸟纲。爬行纲并没有可以鉴定的共有衍征。爬行纲的定义方式包含拥有与缺乏的特征:缺乏羽毛与毛发的羊膜动物。换句话说,亲缘分支分类法学家提出,传统的爬行纲是“非鸟类、也并非哺乳类的羊膜动物”
在1916年,Edwin Stephen Goodrich建立了蜥形纲与兽形纲,用来区别蜥蜴、鸟类与其近亲,以及哺乳类与其已灭绝近亲。两个演化支的心脏与血管差异,以及前脑结构等特征,证实这个分类法。根据Goodrich宣称,蜥形纲与兽形纲共同从一个较早期的原始动物演化而来,名为“Protosauria”,该分类单元也包含某些古生代的两栖类以及早期爬行动物 。
在1956年,David Meredith Seares Watson发现这两个分类单元在爬行动物历史的非常早期就开始分开演化,因此他将“Protosauria”分为蜥形纲与兽形纲。他也分别将鸟类与哺乳类排除在蜥形纲与兽形纲以外。因此根据他的说法,蜥形纲包含:前棱蜥目、始鳄目、米勒古蜥科、龟鳖目、有鳞目、喙头目、鳄目、槽齿目、恐龙、翼龙目、鱼龙目、以及鳍龙超目 。
Watson的分类法补充了Goodrich版本的盲点,但从未广为接受。根据美国古生物学家阿尔弗雷德·罗默(Alfred Sherwood Romer)的著作《 Vertebrate Paleontology 》 ,依照头颅骨的颞颥孔数量与不同位置,爬行纲分为四个亚纲,分别为:
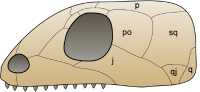
无孔亚纲:没有颞颥孔。
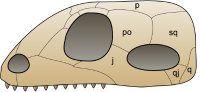
单孔亚纲:一个下方颞颥孔。目前被称为合弓纲。
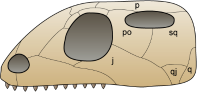
调孔亚纲:一个上方颞颥孔。目前被包含于双孔亚纲之内。
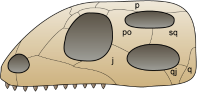
双孔亚纲:两个颞颥孔。
以上四个亚纲,除了单孔亚纲以外,目前都属于蜥形纲。
分类学
无孔亚纲
单孔亚纲
调孔亚纲
双孔亚纲
以下目级列表是根据班顿在2004年的研究 :
羊膜动物
系统发生学
以下简易版的爬行动物演化树 :
演化历史

最早的爬行动物林蜥
最早期的爬行动物,出现于石炭纪晚期,约3亿2000万-3亿1000万年前,演化自迷齿亚纲的爬行形类(Reptiliomorpha) 。林蜥是已知最古老的爬行动物,身长约20到30公分,化石发现于加拿大的新斯科细亚省 。西洛仙蜥( Westlothiana )曾被认为是最早的爬行动物,但目前被认为较接近于两栖类,而非羊膜动物。油页岩蜥与中龙都为最早期的爬行动物之一。最早期的爬行动物生存于石炭纪晚期的沼泽森林,但体型小于同时期的迷齿螈类,例如原水蝎螈( Proterogynrius )、 Cochleosaurus 。石炭纪末期的小型冰河期,使得早期爬行动物有机会成长至较大的体型。
演化支
在最早的爬行动物出现后不久,出现了两个演化支 ,一个是无孔亚纲。无孔亚纲拥有坚硬的头颅骨,没有颞颥孔,仅有与鼻孔、眼睛、脊椎相对应的洞孔 。乌龟被认为是目前仅存的无孔动物,因为它们拥有相同的头颅骨特征;但最近有些科学家认为乌龟是返祖遗传到原始的状态,以增加它们的保护能力 。关于乌龟的起源,目前有副爬行动物的后代、失去颞颥孔的双孔亚纲两派学说 。
另一群演化支是双孔亚纲,头颅骨上有两个颞颥孔,位于眼睛后方。双孔动物进一步分化为两个支系:鳞龙类包含现代蜥蜴、蛇、喙头蜥,可能还有中生代的已灭绝海生爬行动物;主龙类包含现代鳄鱼与鸟类,以及已灭绝的翼龙目与恐龙。
而最早期、具坚硬头颅骨的羊膜动物也演化出另一独立的演化支,称为单孔亚纲。合弓动物的眼睛后方有一对窝孔,可减轻头颅骨重量,并提供颌部肌肉附着点,增加咬合力 。单孔亚纲最后演化为哺乳类,因此被称为似哺乳爬行动物。单孔亚纲过去为爬行纲的一个亚纲,但目前为个别的合弓纲。
二叠纪
在石炭纪末期,合弓类、爬行类动物成为陆地优势动物。迷齿螈类、爬行形类仍然生存在水边,而合弓类盘龙目首先演化至较大的体型,例如基龙与异齿龙。在二叠纪中期,气候多次变迁,造成生态系统的改变,兽孔目取代盘龙目,成为陆地优势动物 。
无孔类爬行动物繁盛于二叠纪。其中,锯齿龙类也演化出较大的体型。大部分无孔类爬行动物在二叠纪-三叠纪灭绝事件灭绝,龟鳖类可能是它们的后代 。
在二叠纪时期,双孔类爬行动物并不繁盛、体型小。但在二叠纪末期演化出两个演化支:主龙形下纲、鳞龙形下纲,最后演化出大部分的现存爬行动物。
中生代
二叠纪末期的二叠纪-三叠纪灭绝事件,造成合弓类动物、无孔类爬行动物的大量灭绝,而主龙形下纲成为陆地优势动物。早期主龙类已具有直立的四足步态,在短期内演化出多种演化支:恐龙、翼龙目、鳄形超目、以及其他三叠纪的主龙类。其中,恐龙是三叠纪后期到白垩纪末期的陆地优势动物群。因此中生代有时被戏称为“恐龙时代”、“爬行动物时代”。在侏罗纪中期,兽脚亚目恐龙演化出许多有羽毛恐龙,更进一步演化出鸟类 。
相对于主龙形下纲,鳞龙形下纲则可能演化出多群海生爬行动物:楯齿龙目、幻龙目、蛇颈龙目、沧龙科;鱼龙类可能演化自更原始的双孔类爬行动物 。鳞龙形下纲也演化出多种陆栖小型爬行动物,例如:喙头蜥、蜥蜴、蛇、蚓蜥。
在恐龙的竞争压力下,兽孔目演化出体型小、高代谢率的物种,并在侏罗纪晚期演化出哺乳动物。
新生代
白垩纪末期的白垩纪-第三纪灭绝事件,使恐龙、翼龙目、大部分海生爬行动物、大部分鳄形类灭绝,而鸟类、哺乳动物在新生代再次繁盛、多样化,因此新生代被戏称为“哺乳动物时代”。只有龟鳖类、喙头蜥、蜥蜴、蛇、蚓蜥、鳄鱼继续存活到现代,主要生存于热带与副热带地区。现存爬行动物大约有8,200个种,其中半数属于蛇。
生物学
循环系统

巨蜥的红外线图像
大部分的现存爬行动物具有闭合的循环系统,它们具有三腔室心脏,由两个心房与一个心室所构成,心室的分割方式并不一致。它们通常只有一对大主动脉。当它们的血液流经三腔室心脏时,含氧血与缺氧血只有少量混合。但是,血液可改变流通方式,缺氧血可流向身体,含氧血可流向肺脏,使爬行动物的体温调节更有效率,尤其是水生物种 。
鳄鱼具有四腔室心脏,可以在水中以三腔室心脏运作 。某些蛇类与蜥蜴(例如巨蜥与蟒蛇),具有三腔室心脏,但可以四腔室心脏方式运作。因为它们心脏的皮瓣可在心动周期时,随者扩张、收缩隔开心室。某些喙头蜥可借由皮瓣,产生类似哺乳动物与鸟类的心脏运动 。
呼吸系统
所有的爬行动物都用肺呼吸。水生乌龟发展出具渗透性的皮肤,某些爬行动物可用泄殖腔来增加气体交换的面积(Orenstein, 2001)。即使具有这些构造,它们仍需要肺来完成呼吸作用。在各个主要的爬行动物种类之间,完成肺部换气的方式并不相同。有鳞目爬行动物的肺部几乎只借由轴肌换气,而这个肌肉也同时用于运动。因为这个限制,大部分的有鳞目爬行动物必须在激烈的移动中停止呼吸。但是,有些爬行动物找到了解决方法。巨蜥以及一些其他的蜥蜴物种使用一种使用到喉咙肌肉的方法来补足原本的“轴呼吸”。这使得这些动物可以在激烈运动中为肺部完全充气,所以可以持续进行有氧呼吸很长一段时间。据了解双领蜥具有原始横膈膜可将胸腔和腹腔分开来。即使此膜不能动,它还是能借着将内脏的重量和肺分开而增加肺的吸气量。鳄鱼其实拥有和哺乳类动物相似的有肌肉横膈膜。不同的是,鳄鱼的横膈膜将耻骨拉回(部分的骨盆,在鳄鱼中能移动),从而使肝下降,以腾出空间使肺能胀大。这种横膈膜式的机制被称作〝肝活塞〞。
关于海龟和陆龟如何呼吸已经被研究了很多次。迄今为止,只有少数物种已经被深入研究到足以了解乌龟如何呼吸。其结果表明,海龟及陆龟分别找到了不同的方法来解决这个问题。但问题是,大部分的龟壳是僵硬的,且不能像其他羊膜动物使用的膨胀和收缩来为肺部换气。一些乌龟,如印度鳖( lissemys punctata )有一层肌肉包覆着肺部。当它收缩时,乌龟可以呼气;在静止时,乌龟可以收回四肢进到体腔内,并强制使空出肺部的。而当乌龟伸出其四肢时肺部内的压力会减少,乌龟因此可以空气。乌龟的肺连接到壳内的上方,而肺的底部附着在其余内脏(由结缔组织连接)。借由一系列特殊的肌肉(大约相当于一个横膈膜),因为许多这些肌肉的附着点都在他们的前肢上(事实上,很多的肌肉在收缩时扩张到四肢的袋子里),乌龟能够推动他们的内脏向上或向下,造成有效的呼吸。现今已研究了三个物种在运动期间的呼吸,并且显示了它们之间不同的模式。当成年雌性绿蠵龟沿着要产卵的海滩爬行时并不呼吸。他们在陆上运动时屏住呼吸,只有在停顿休息时呼吸。北美的箱龟运动期间还是继续呼吸,而且其换气的周期并不与肢体动作相关。它们可能是利用其腹部肌肉在运动中呼吸。最后被研究的物种是红耳龟。它们也在运动中呼吸,但他们在运动中的呼吸比在运动间停顿时的呼吸还少,显示肢体运动和呼吸器具之间可能有机械干扰。箱龟也被观察到在完全密封的壳内呼吸。
骨骼系统
爬行动物的骨骼系统大多数由硬骨组成,骨骼的骨化程度高,很少保留软骨部分。
大部分的爬行动物缺乏次生颚,所以当它们进食时,无法同时呼吸。鳄鱼已发展出骨质次生颚,使它们可在半隐没至水中时持续呼吸,并防止嘴中的猎物挣扎时,伤及脑部。石龙子科也演化出骨质次生颚。
排泄系统
爬行动物的排泄系统主要借由两颗肾脏。双孔动物所排泄的主要含氮废物是尿酸;泽龟龟主要排泄尿素,陆龟如苏卡达、星龟排尿酸,而部分则排尿液,类似哺乳类。不像哺乳类与鸟类的肾脏,爬行动物的肾脏不能够制造尿液,尿液可以储藏更多的身体废物。这是因为它们缺乏鸟类与哺乳类所拥有的肾元,称为亨利氏环(Loop of Henle)。因此许多爬行动物使用它们的结肠与排泄腔来再度吸收水分。有些爬行动物甚至可利用膀胱来取得水分。有些爬行动物的鼻部与舌部具有盐腺,可以储存额外的盐分。
神经系统
爬行动物与两栖类的脑部具有相同的基本部分,但它们的大脑与小脑稍大。爬行动物的感觉器官多发展良好,除了少部分物种,例如蛇缺乏外耳,但仍具有中耳与内耳。爬行动物具有12对脑神经[1]。
繁衍
除了陆龟与海龟以外,大部分的雄性爬行动物具有成对的管状性器官,称为半。陆龟与海龟则具有单一。所有的龟鳖目皆为卵生动物,而某些蜥蜴与蛇是卵胎生或胎生动物。爬行动物借由泄殖腔来交配、繁衍;泄殖腔位于尾巴基部,可用来排泄与繁殖。
对于某些有鳞目物种而言,雌性个体若达到一定的数量,它们便能自行复制出单性染色体。这种无性繁殖方式称为孤雌生殖(Parthenogenesis),目前已有六科蜥蜴,以及一种蛇,被确认具有这种无性繁殖能力,例如:数种壁虎、美洲蜥蜴科(尤其是 Aspidocelis )、以及蜥蜴科。被封闭的科莫多龙(巨蜥科)也具有孤雌生殖能力。
除了以上物种以外,变色龙、飞蜥科、夜蜥蜴科、以及嵴尾蜥科,也被推测具有孤雌生殖能力。
爬行动物的蛋,外部是钙质蛋壳或皮革,覆盖者内部的羊膜、羊膜囊、以及尿囊。
参考文献

盾臂龟原居撒哈拉沙漠南部
^ Benton, Michael J.Vertebrate Palaeontology3rd. Oxford: Blackwell Science Ltd. 2005. ISBN 978-0-632-05637-8.
^ Benton, Michael J. Vertebrate Palaeontology 4th. Oxford: Blackwell Science Ltd. 2014. ISBN 978-0-632-05637-8.
^ Linnaeus, Carolus.Systema naturae per regna tria naturae :secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis.10th. Holmiae (Laurentii Salvii). 1758 [ 2008-09-22 ] (Latin) . 引文格式1维护:未识别语文类型 (link)
^ Laurenti, J.N. (1768): Specimen Medicum, Exhibens Synopsin Reptilium Emendatam cum Experimentis circa Venena .Facsimile, showing the mixed composition of his Reptilia
^ Encyclopaedia Britannica, 9th ed. (1878).original text
^ Latreielle, P.A. (1804): Nouveau Dictionnaire à Histoire Naturelle, xxiv., cited in Latreille"s Familles naturelles du règne animal, exposés succinctement et dans un ordre analytique , 1825
^ Tudge, p.85
^ Goodrich, E.S. On the classification of the Reptilia. Proceedings of the Royal Society of London. 1916, 89B : 261–276.
^ Watson, D.M.S. On Millerosaurus and the early history of the sauropsid reptiles. Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences. 1957, 240 (673): 325–400.
^ Romer, A.S. Vertebrate Paleontology. University of Chicago Press. 1933. , 3rd ed., 1966.
^ Benton, Michael J. Vertebrate Paleontology 3rd ed. Oxford: Blackwell Science Ltd. 2004. ISBN 0632056371. 引文格式1维护:冗余文本 (link)
^ Laurin, M. and Gauthier, J.A. (1996). "Amniota. Mammals, reptiles (turtles, lizards, Sphenodon, crocodiles, birds) and their extinct relatives." Version 01 January 1996. /Amniota/14990/1996.01.01in The Tree of Life Web Project, /
^13.013.1 Laurin, M.; Reisz, R. R. A reevaluation of early amniote phylogeny. Zoological Journal of the Linnean Society. 1995, 113 : 165–223. doi:10.1111/j.1096-3642.1995.tb00932.x . (abstract)
^ Falcon-Lang, H.J., Benton, M.J. & Stimson, M. (2007): Ecology of early reptiles inferred from Lower Pennsylvanian trackways. Journal of the Geological Society , London, 164; no. 6; pp 1113-1118.article
^ Palmer, D. (编). The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London: Marshall Editions. 1999: 62. ISBN 1-84028-152-9.
^ van Tuninen, M. & Hadly, E.A. (2004): Error in Estimation of Rate and Time Inferred from the Early Amniote Fossil Record and Avian Molecular Clocks. Journal of Mulecular Biology , no 59: pp 267-276PDF
^17.017.1 Romer, A.S. & T.S. Parsons. 1977. The Vertebrate Body. 5th ed. Saunders, Philadelphia. (6th ed. 1985)
^ Benton, M. J. Vertebrate Paleontology 2nd. London: Blackwell Science Ltd. 2000. ISBN 0632056142. , 3rd ed. 2004 ISBN 978-0-632-05637-8
^ Zardoya, R.; Meyer, A.Complete mitochondrial genome suggests diapsid affinities of turtles. Proc Natl Acad Sci U S A. 1998, 95 (24): 14226–14231. ISSN 0027-8424 . PMID 9826682 . doi:10.1073/pnas.95.24.14226 .
^ Rieppel, O.; deBraga, M. Turtles as diapsid reptiles. Nature. 1996, 384 : 453–455. doi:10.1038/384453a0 .
^ Colbert, E.H. & Morales, M. (2001): Colbert"s Evolution of the Vertebrates: A History of the Backboned Animals Through Time . 4th edition. John Wiley & Sons, Inc, New York - ISBN 978-0-471-38461-8.
^ Colbert, E.H. & Morales, M. (2001): Colbert"s Evolution of the Vertebrates: A History of the Backboned Animals Through Time . 4th edition. John Wiley & Sons, Inc, New York - ISBN 978-0-471-38461-8.
^ Colbert, E.H. & Morales, M. (2001): Colbert"s Evolution of the Vertebrates: A History of the Backboned Animals Through Time . 4th edition. John Wiley & Sons, Inc, New York - ISBN 978-0-471-38461-8.
^ Gauthier J. A. (1994): The diversification of the amniotes . In: D. R. Prothero and R. M. Schoch (ed.) Major Features of Vertebrate Evolution: 129-159. Knoxville, Tennessee: The Paleontological Society.
^ Hicks, James. The Physiological and Evolutionary Significance of Cardiovascular Shunting Patterns in Reptiles. News in Physiological Sciences. 2002, 17 : 241–245.
^ Axelsson, Michael; Craig E. Franklin. From anatomy to angioscopy: 164 years of crocodilian cardiovascular research, recent advances, and speculations.. Comparative Biochemistry and Physiology A. 1997, 188 (1): 51–62.
^ Wang, Tobias; Altimiras, Jordi; Klein, Wilfried; Axelsson, Michael. Ventricular haemodynamics in Python molurus: separation of pulmonary and systemic pressures. The Journal of Experimental Biology. 2003, 206 : 4242–4245. PMID 14581594 . doi:10.1242/jeb.00681 .
参见
爬行动物分类表
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。




- 有价值
- 一般般
- 没价值









24小时热门







推荐阅读


关于我们

APP下载


