





-
 QQ空间
QQ空间
-
 QQ好友
QQ好友
-
 微信好友
微信好友
-
 新浪微博
新浪微博
转运核糖核酸
tRNA的发现
1955年Zamecnik认为标记的ATP可能参与RNA的生物合成。于是他将 C标记的ATP与微粒体(Microsome)和细胞抽提液的可溶性部分一起保温后,发现RNA居然也被标记了。他有点怀疑。可是,当他将 C标记的氨基酸与微粒体和可溶性部分在同样条件下保温后,他惊奇地发现,与RNA合成无关的 C氨基酸也标记了RNA,而且更意想不到的是 C标记的RNA不是核糖体的大分子RNA,而是可溶性部分中的小分子RNA。进一步,仅将可溶性部分与 C标记的氨基酸和ATP一起保温,则这种 C标记的氨基酸仍能与其中的小分子RNA结合。因此,这种可溶性部分中的小分子RNA被称为称sRNA(soluble RNA)。1956年Watson曾访问Zamecnik实验室,并对他们说,1955年Crick已经提出过“适配子”的设想。后来,这种 sRNA被命名为tRNA。
tRNA的结构
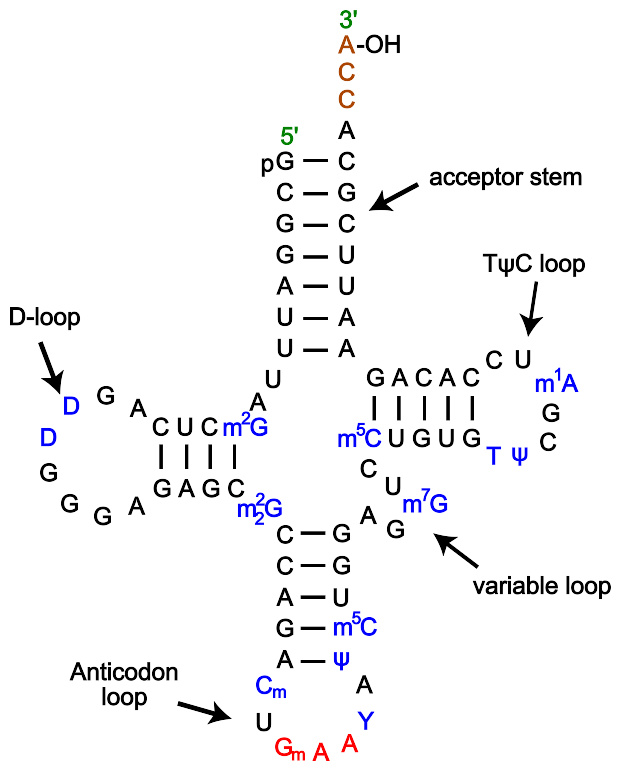
来自酵母转运核糖核酸苯丙氨酸次级三叶草结构
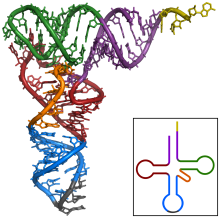
tRNA的结构
tRNA为74~95个碱基的小片段RNA链,会折叠成苜蓿叶状的二级结构,呈三叶草形,它由氨基酸臂、二氢尿嘧啶环、反密码环、额外环和TΨC环五部分组成。
tRNA有一级结构(5"到3"的核苷酸方向),二级结构(通常显示为三叶草结构)和三级结构(所有的tRNA具有类似L-形的三维结构,允许它们与核糖体的P、A位点结合)。
特色
5"端磷酸。
受体臂(accept stem,也被称作 amino acid stem )是一个7个碱基长的臂,其中包含5"端,与有3"端羟基(OH,能结合氨基酸于其上)的3"端。受体臂有可能含有非Watson-Crick所发现的碱基对。
CCA尾(CCA tail)是tRNA分子3"端的CCA序列,在翻译时,帮助酶识别tRNA。
D臂(D arm)是在一个环(D loop)的端部4个碱基的臂,通常含有二氢尿嘧啶(dihydrouridine)。
反密码子臂(anticodon arm)有5个碱基,包括反密码子(anticodon)。每一tRNA包括一个特异的三联反密码子序列,能够与氨基酸的一个或者多个密码子匹配。例如赖氨酸(lysine)的密码子之一是AAA,相应的tRNA的反密码子可能是UUU(一些反密码子可以与多于一个的密码子匹配被称为“摆动”)。
T臂(T arm)是5个碱基的茎,包括序列TψC。
修饰碱基(Modified bases)是tRNA中的一些不常见的碱基,如腺嘌呤、鸟嘌呤、胞嘧啶和尿嘧啶的修饰形式。
氨酰化
氨酰化(Aminoacylation)是添加一个氨酰基团到化合物的过程。
在氨酰tRNA合成酶(aminoacyl tRNA synthetase)的作用下,tRNA与特异的氨基酸进行氨酰化反应(aminoacylated)。对于一种氨基酸而言,尽管可能有多种 tRNA和多种反密码子,但是通常只有一种氨酰tRNA合成酶。合成酶对合适的tRNA的识别不仅仅是反密码子,受体臂也起了显著的作用。
反应:
氨基酸 + ATP →氨基酰- AMP + PPi
氨基酰-AMP + tRNA →氨基酰 - tRNA + AMP
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读
知识互答

关于我们

APP下载



{{item.time}} {{item.replyListShow ? '收起' : '展开'}}评论 {{curReplyId == item.id ? '取消回复' : '回复'}}