





-
 QQ空间
QQ空间
-
 QQ好友
QQ好友
-
 微信好友
微信好友
-
 新浪微博
新浪微博
螯肢亚门
特征
节及表皮
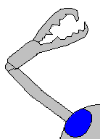
广翅鲎的螫肢。

蜘蛛的螫肢,可以让螫爪完全向外翻。
螫肢亚门的身体及肢体有节,并由甲壳素及蛋白质组成的表皮覆盖。其头部由几节组成,这些节于胚胎发育时就已经融合在一起。体腔非常细小,血腔连接至管状的心脏。它们的身体分为两个体区,最前端的头与及后方的腹部。但是蜱螨亚纲的头及腹部并没有明显的分野。
螫肢亚门的头于胚胎时期透过融合带有眼睛的口前叶和第二至第七节生成,第一节于胚胎发育期间会消失。第二节有一对螫肢,第三节有一对须肢,很多亚类都是以此来感官的,其余的四节都有脚。较原始形态的口前叶两侧有一对复眼,中央有四只单眼。口器是在第二及第三节之间。
腹部包含12或更少的节数,头七节为前腹部,后五节为后腹部,以尾节或尖刺终结。现今螫肢亚门已经失去了前腹部附肢,或是已经大幅演化,如蜘蛛的喷丝板或鲎的鳃。
螫肢亚门的身体及附肢都由粗糙的表皮所覆盖,由于表皮不能伸展,它们需要脱皮来成长。
螫肢及须肢
螫肢亚门的螫肢及须肢在形态及功能上有很大分野,唯一相同的就是其位置:螫肢出于第竹二节,位于口器前;而须肢来自第三节,位于口器之后。
螫肢一般分三节,第三节上有螫爪。不过,蜘蛛的螫肢只有两节,于第二节有螫爪,不使用时螫爪可以收起。螫肢的大小也各有不同:广翅鲎的螫爪很大,而蝎子的则很细小。
大部分螫肢亚门的须肢相对较为细小及用作感官。不过雄性蜘蛛的须肢端呈球状,可以作为针筒将精子注入雌性的生殖口,而蝎子的则有大钳用来捕捉猎物。
体腔及循环系统
螫肢亚门的体腔非常细小,在生殖系统及排泄系统之外只余少许空间。主体腔是将血液带到全身的血腔,管状的心脏会将后部的血液向前供血。虽然动脉会将血液带到特定的身体部位,但末端是开放的,并没有接合至静脉,是为开放式的循环系统。
呼吸系统
螫肢亚门的呼吸系统会就亚类所生存的环境而有所不同。现存陆生的螫肢亚门一般都有书肺及气管,书肺会经血液供应氧及移除废气,而气管可以不用经血液而发挥同样功能。鲎是水生的,故在类同位置有书鳃。已灭绝的广翅鲎亚纲估计是有鳃的,但其化石证明却很混淆。但是奥陶纪晚期Onychopterella的化石似乎有三对书鳃,外观非常像蝎子书肺。
觅食及消化
大部分螫肢亚门的食道太窄,不足以容纳固体食物。所有蝎子及差不多所有蜘蛛都是掠食者,并会预先在口前腔用螫肢处理猎物。不过有一种蜘蛛是草食性的,并有很多是会以花蜜及花粉来补充其营养。大部分蜱螨亚纲都是吸血的寄生虫,但也有掠食性、草食性及食腐性的亚类。所有蜱螨亚纲都有可收缩的螫肢、须肢及部分外骨骼,组成口前腔来预先处理食物。
盲蛛目是少数现存螫肢亚门可以吃固体食物,当中也有掠食性、草食性及食腐性的。鲎有独特的消化系统可以处理固体食物。它们脚端的爪会将细小的无脊椎动物抓入最后肢至口器间的食物沟,脚基位置有锯齿状的颚基可以磨碎食物,并将之放入口内。这种方式相信是最古老的节肢动物的觅食方法。
排泄
鲎会将氮废物转为氨,再经鳃排出。其他的废物会成为粪便由肛门排出,另外亦有肾管遣滤出尿液。氨是有毒的,故它们需要大量水份来稀释。大部分陆生的螫肢亚门都不能消耗如此大量的水分,故会将氮转为其他化合物,并以固体形式排出。排泄可以由肾管及马氏管进行。这些管都是从血液中过滤废物,并以固体形态排到后肠。
神经系统
螫肢亚门的神经系统都是基于节肢动物的一对标准神经索,每节的神经索上都有神经节。它们的脑部是神经节的融合,位于口器之后。不过,由于螫肢二门的第一节会消失,所以它们的脑部只包含一对口前神经节,而不像其他昆虫有两对。其他神经节融入至脑部的方式各有不同。鲎的脑部包含了所有头及首两腹节的神经节,而其他腹节的神经节则保持分离。除了蝎子以外,大部分现存蛛形纲的所有神经节都在头融合成单一团,而腹部则完全没有神经节。古疣亚目腹部及头胸后部的神经节并没有融合,而蝎子头的神经节融合在一起,但腹部的则保持分离。
感觉
螫肢亚门的表皮阻隔了外界的资讯,只透过传感器或神经系统的连接来获得感觉。蜘蛛及其他节肢动物都改进了其表皮成一系列的传感器。绒毛是触觉及振动的传感器,由强烈的接触至非常弱的气流也可以感知。化学传感器提供了与味觉及嗅觉一样的功用。
现存螫肢亚门拥有复眼及单眼,复眼位于头部两侧,单眼位于中央。鲎的眼睛可以在没有形成影像的情况下侦测运动。蝇虎科有非常阔的视觉,其主眼比蜻蜓的准确多十倍。
繁殖

雌性Compsobuthus werneri蝎子,背上白色的是幼体。
螫肢亚门的育婴程度各有不同。蝎子会将幼体放在背上直至它们第一次脱壳,而一些物种就会一直留在母亲身边。一些蜘蛛会照顾其幼体,如狼蛛科的幼体会抓着母亲的绒毛,而母亲亦会报以猎物作为回应。
演化历史
化石纪录
螫肢亚门的化石纪录有很大的断层,这是由于它们的外骨骼是有机的,很难作为化石被保存下来,除非是在特别适合保存软组织的地层才能有所发现。伯吉斯页岩中属于5亿500万年前的多须虫及西德尼虫都被分类到螫肢亚门中,因为前者有体区分段,而后者则有像剑尾目的附肢。支序分类学分析却认为它们有更多的特征并不属于螫肢亚门。5亿2500万年前寒武纪早期抚仙湖虫是否属于螫肢亚门激起了争论。另一个寒武纪的科氏惊异虫原先被分类到Aglaspidida中,但有可能是属于广翅鲎亚纲。若这些任何一种是与螫肢亚门有关,化石断层最少也有4300万年。
Palaeotarbus jerami是已知最古老的蛛形纲。
已知最古老的广翅鲎亚纲可以追溯至4亿3600万至4亿2800万年前的志留纪,但于2008年发现了属于4亿4500万年前奥陶纪晚期的标本。
已知最古老的蛛形纲是Palaeotarbus jerami,源自4亿2000万年前的志留纪。它们拥有三角形的头及有节的腹部,有八对脚及一对须肢。
3亿3600万年前泥盆纪的Attercopus fimbriunguis是最早拥有生产丝器官的,但却没有喷丝板,故并非真正的蜘蛛。有几种石炭纪的蜘蛛都是属于中突蛛亚目。
志留纪晚期的Proscorpius被分类为蝎子,但明显与现今的蝎子有所分别。它们是完全水生的,用鳃呼吸。其口器位于头部底,接近第一对脚之间,有点像已灭绝的广翅鲎亚目及现存的鲎。4亿200万年前泥盆纪早期的陆生蝎子化石拥有书肺。
与其他的关系
螫肢亚门近期的种系发生。
传统上螫肢亚门的系统发生。
传统上螫肢亚门与其他主要的节肢动物门较为疏远。但近期的分子种系发生学及对节肢动物的神经系统在胚胎时期的发育研究,显示螫肢亚门与多足纲较为接近。不过,研究结果只是针对现存的节肢动物,若结合已灭绝的动物(如三叶虫),结果可能会重返传统的观点上。
主要亚类
蛛形纲的演化树。
一般认同螫肢亚门包含了蛛形纲、剑尾目及广翅鲎亚纲。已灭绝的Chasmataspida是广翅鲎亚目下的亚类。海蜘蛛纲传统上分类在螫肢亚门之内,但有些特征显示它们是演化自最古老的节肢动物。
不过自19世纪,螫肢亚门演化树内的关系就已经被受争议。2002年综合现今螫肢亚门的RNA特征及现今和已灭绝的螫肢亚门的解剖特征后,为低层亚类提供了可靠的结果,但在高阶的亚类中并不稳定。2007年的研究提供了右侧的演化树,但却仍有一些不明处。
蝎子的分类位置特别受到争论。一些早期被认为是蝎子的化石,如志留纪晚期的Proscorpius却有鳃和是水生的,口器的位置就像广翅鲎亚目及鲎一样。这可以有三个选择:将这些水生的化石分类到其他地方;或接受蝎子并非单系群,而是有水生及陆生的;或是将蝎子分类到较为接近广翅鲎亚目的位置,而疏于蜘蛛及其他蛛形目,蝎子因而并非属于蛛形目,而蛛形目亦非单系群。
多样性
虽然不及昆虫,螫肢亚门是最多样化的动物之一,其下有超过77000个现存物种。估计仍有13万种蜘蛛及50万种螨未被描述。虽然最早的螫肢亚门及现存的海蜘蛛及剑尾目都是水生及呼吸溶氧,其他的都是呼吸空气的,除了一些蜘蛛可以将蜘蛛网织成潜水钟的形状进入水中生活。大部分现存的螫肢亚门都是掠食者,主要掠食细小的无脊椎动物。其他很多都是寄生、草食性、食腐性及食碎屑性的。
与人类的关系
以往美国原住民是会吃鲎,用其尾棘制作矛头,壳用来舀水。不过他们已经放弃了将鲎作为牲畜食物,因为发现会令牲畜肉质的味道变坏。鲎的血液含有鲎阿米巴样细胞溶解物(Limulus Amebocyte Lysate)可以用作细菌存在的试剂,以及用来侦测脑膜炎及一些癌症。
在柬埔寨及委内瑞拉南部会煮食捕鸟蛛科,并视为是一种美食。蜘蛛毒素对环境的污染较传统的杀虫剂为低,因只会对昆虫(而不是脊椎动物)起作用。蜘蛛毒素也正研究用作医药来治疗心律不整、阿兹海默病、中风及勃起功能障碍。由于蜘蛛丝轻巧而坚韧,基因工程师尝试利用羊奶及叶子来制作蜘蛛丝。于20世纪就有约100宗被蜘蛛咬死的个案,相对于水母刺死的1500宗为小。
蝎子刺螫亦是相当危险的,例如在墨西哥每年就约有1000宗死亡个案,但在发展较先进的美国每年就只有1宗。大部分个案都是因人类误闯蝎子巢穴而发生的。蝎子毒素也正被开发来作为医药用途,包括治疗脑癌及骨骼病症。
蜱是寄生的,一些会传播引发疾病的微生物及寄生虫,一些的口水更会直接引起蜱瘫痪。
一些螨也会侵扰人类,它们咬或钻入皮肤下而引起痕痒。那些侵扰啮齿目等动物的物种在失去主体时也会影响人类。有三种螨会对蜜蜂造成危害,其中一种的瓦螨是养蜂人的劲敌。螨会引起多种过敏症,包括过敏性鼻炎、哮喘及湿疹,与及加剧异位性皮肤炎。它们也是害虫,但掠食性的螨可以控制它们。
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。




- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读

知识互答

关于我们

APP下载



{{item.time}} {{item.replyListShow ? '收起' : '展开'}}评论 {{curReplyId == item.id ? '取消回复' : '回复'}}