





-
 QQ空间
QQ空间
-
 QQ好友
QQ好友
-
 微信好友
微信好友
-
 新浪微博
新浪微博
染色体
发现历程
1879年,德国生物学家华尔瑟·弗莱明(Walther Flemming)称这种丝状体为染色体( 英语: chromosome ;希腊语: chroma =颜色,希腊语: soma =体),意即可染色的小体,并猜测染色体与遗传有关。1902年,博韦里(T·Boveri)和萨顿(W·S·Sutton)指出,染色体在细胞分裂中的行为与孟德尔的遗传因子平行:两者在体细胞中都成对存在,而在生殖细胞中则是单独存在的;成对的染色体或遗传因子在细胞减数分裂时彼此分离,进入不同的子细胞中,不同对的染色体或遗传因子可以自由组合。因此,博韦里和萨顿认为,染色体很可能是遗传因子的载体。
结构

染色体结构
核小体是染色体结构的最基本单位。核小体的核心是由4对组织蛋白(H2A、H2B、H3和H4)各两个分子构成的扁球状8聚体。现在我们知道,脱氧核糖核酸分子具有典型的双螺旋结构,一个脱氧核糖核酸分子就像是一条长长的双螺旋的飘带。一条染色体有一个脱氧核糖核酸分子。脱氧核糖核酸双螺旋依次在每个组蛋白8聚体分子的表面盘绕约1.75圈,其长度相当于140个碱基对。组蛋白8聚体与其表面上盘绕的脱氧核糖核酸分子共同构成核小体。在相邻的两个核小体之间,有长约50~60个碱基对的脱氧核糖核酸连接线。在相邻的连接线之间结合着一个第5种组蛋白(H1)的分子。密集成串的核小体形成了核质中的100埃左右的纤维,这就是染色体的“一级结构”。在这里,脱氧核糖核酸分子大约被压缩了7倍。
染色体的一级结构经螺旋化形成中空的线状体,称为 螺线体或称核丝 ( 英语 : Solenoid (DNA) ) ,这是染色体的“二级结构”,其外径约300埃,内径100埃,相邻螺旋间距为110埃。螺丝体的每一周螺旋包括6个核小体,因此脱氧核糖核酸的长度在这个等级上又被再压缩了6倍。
300埃左右的螺线体(二级结构)再进一步螺旋化,形成直径为0.4微米的筒状体,称为超螺旋体。这就是染色体的“三级结构”。到这里,脱氧核糖核酸又再被压缩了40倍。超螺旋体进一步折叠盘绕后,形成 染色单体 —染色体的“四级结构”。两条染色单体组成一条染色体。到这里,脱氧核糖核酸的长度又再被压缩了5倍。从染色体的一级结构到四级结构,脱氧核糖核酸分子一共被压缩了7×6×40×5=8400倍。例如,人的染色体中脱氧核糖核酸分子伸展开来的长度平均约为几个厘米,而染色体被压缩到只有几个微米长。
现代关于染色体超微结构的概念
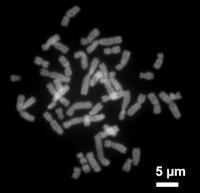
电子显微镜下的人类染色体。
染色体的超微结构显示染色体是由直径仅100埃(Å)的去氧核糖核酸-组蛋白高度螺旋化的纤维所组成。每一条染色单体可看作一条双螺旋的脱氧核糖核酸分子。有丝分裂间期时,解螺旋而形成无限伸展的细丝,此时不易为染料所着色,光镜下呈无定形物质,称之为染色质。有丝分裂时脱氧核糖核酸高度螺旋化而呈现特定的形态,此时易为碱性染料着色,称之为染色体。
1970年后陆续问世的各种显带技术对染色体的识别作出了很大贡献。中期染色体经过DNA变性、胰酶消化或荧光染色等处理,可出现沿纵轴排列的明暗相间的带纹。按照染色体上特征性的标志可将每一个臂从内到外分为若干区,每个区又可分为若干条带,每条带又再分为若干个亚带,例如“9q34.1”即表示9号染色体长臂第3区第4条带的第1个亚带。由于每条染色体带纹的数目和宽度是相对恒定的,根据带型的不同可识别每条染色体及其片段。
1980年代以来根据脱氧核糖核酸双链互补的原理,应用已知序列的DNA探针进行荧光原位杂交(Fluorescence in situ hybridization,FISH)可以识别整条染色体、染色体的1个臂、1条带甚至一个基因,因而大大提高了染色体识别的准确性和敏感性。染色体是遗传物质—基因的载体,控制人类形态、生理和生化等特征的结构基因呈直线排列在染色体上。由此可见,染色体和基因二者密切相关,染色体的任何改变必然导致基因的异常。
2000年6月26日人类基因组计划(HGP)已宣布完成人类基因组序列框架图。2001年2月12日HGP和塞雷拉公司公布了人类基因组图谱和初步分析结果。人类基因组共有3~3.5万个基因,而不是以往认为的10万个。
型态
染色体在细胞分裂之前才形成。在细胞的代谢期或间期,染色体分散成一级结构或伸展开的脱氧核糖核酸分子,组成细胞核内的染色质或核质。
染色体的形态以中期时最为典型。每条染色体由两条染色单体组成,中间狭窄处称为着丝点(centromere),又称主缢痕,它将染色体分为短臂(p)和长臂(q)。按着丝粒位置的不同,人类染色体可分为 中着丝粒染色体 、 亚中着丝粒染色体 和 近端着丝粒染色体 等3种类型。近端着丝粒染色体的短臂末端有一个叫做随体的结构,它呈圆球形,中间以细丝与短臂相连。有的染色体长臂上还可看到另一些较小的狭窄区,称为次缢痕。染色体臂的末端存在着一种叫做端粒(telomere)的结构,它有保持染色体完整性的功能。
各种生物体细胞核内染色体数目
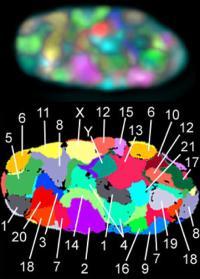
在成人纤维细胞上的23种(21+X+Y)人类染色体。
真核生物
真核细胞中细胞核外的其他染色体(例如线粒体内的小染色体或类质粒小染色体)的数量是不固定的,可能数以千计。
进行无性生殖的物种的所有细胞中只有一套染色体,这一套染色体在所有体细胞中都是相同的。
进行有性生殖的物种的体细胞中有两套染色体,一套来自父方;一套来自母方。而生殖细胞只有一套染色体,这一套染色体来自于具两套染色体精原细胞或卵母细胞的减数分裂。减数分裂进行时,同大染色体(一对成对的染色体)可能会染色体互换,由此产生与父母方的都不完全一样,子代不是完全继承父方或母方的新染色体。
某些生物是多倍体,体细胞有三套甚至更多套染色体。
巴尔氏体
巴尔氏体(英语:Barr body,也译作巴氏小体或巴尔小体),得名自其发现者穆雷·巴尔,是具有两个以上X染色体(或Z染色体)的细胞中,为了避免该染色体上基因的过度表现,而将其中一条X染色体或Z染色体去活化而成的紧密结构。X染色体去活化机制由“Xist基因”控制,该基因可使被去活化的X染色体受到较高度的DNA甲基化与较低度的[组蛋白胺酸化](histone acetylation),使其成为一种异染色质,即巴尔氏体。
巴尔氏体上除了伪体染色体区(pseudoautosomal region)的基因仍可表现外,其余片段的基因都受到抑制,这种现象称为基因沉默(gene silencing)。巴尔氏体产生的目的和避免雌性的X染色体(或雄性的Z染色体)剂量过多有关,造成雌性身体由关闭不同X染色体的两群不同的细胞组成,称为镶嵌体(mosaic),可以产生两种不同的表现型,这也是花猫有三种毛色的原因。
染色体数目的变异
正常情况下,个生物体内,含有一定数量的染色体(见上表),而且是成对出现(2n)。而多倍体生物的细胞内具有超过两套以上的染色体。三倍体的生成,可能是在形成配子的减数分裂的过程中,染色体发生了无分离(nondisjunction)现象而出现了2n的配子,故受精后产生三倍体的子代。四倍体的形成,可能是在2n的合子再行有丝分裂时,复制完成的染色体发生了无分离,导致产生含有四套染色体的子代。多倍体的果实通常都较巨大。
外部链接
参见
遗传删除
染色体遗传算法
遗传算法
染色体双中节
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。



- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读

知识互答

关于我们

APP下载



{{item.time}} {{item.replyListShow ? '收起' : '展开'}}评论 {{curReplyId == item.id ? '取消回复' : '回复'}}