






白垩纪-第三纪灭绝事件
灭绝模式
虽然白垩纪﹣第三纪灭绝事件造成许多物种灭绝,但不同的演化支,或是各个演化支内部,呈现出明显差异的灭绝程度。由于大气层中的微粒遮辟了阳光,减少抵达地表的太阳能,依赖光合作用的生物衰退或灭绝。在白垩纪晚期,食物链底层是由依赖光合作用的生物构成,例如浮游植物与陆地植物,如同现今的状况。证据显示,草食性动物因所依赖的植物衰退,而数量减少;同样地,顶级掠食者(例如暴龙)也接连受到影响。
颗石藻与软体动物(包含菊石亚纲、厚壳蛤、水生蜗牛、蚌),还有以上述硬壳动物维生的动物,在这次灭绝事件中灭亡,或遭受严重打击。例如,沧龙类被认为以菊石为食,这群海生爬行动物在白垩纪﹣第三纪灭绝事件中灭亡。
杂食性、食虫性、以及食腐动物在这次灭绝事件中存活,可能因为它们的食性较多变化。白垩纪末期似乎没有完全的草食性或肉食性哺乳动物。哺乳动物与鸟类借由以昆虫、蚯蚓、蜗牛等动物为食,而在K-T事件中存活,而这些动物则以死亡的植物与动物为食。科学家假设,这些生物以生物的有机碎屑为生,因此得以在这次植物群崩溃的灭绝事件存活。
在河流生物群落中,只有少数动物灭亡;因为河流生物群落多以自陆地冲刷下来的生物有机碎屑为生,较少直接以活的植物为生。 海洋也有类似的状况,但较为复杂。生存在浮游带的动物,所受到的影响远比生存在海床的动物还大。生存在浮游带的动物几乎以活的浮游植物为生,而生存在海床的动物,则以生物的有机碎屑为食,或者可转换成以生物的有机碎屑为食。
在这次灭绝事件存活下来的生物中,最大型的陆地动物是鳄鱼与离龙目,是半水生动物,并可以生物碎屑为生。现代鳄鱼可以食腐为生,并可长达数月未进食;幼年鳄鱼的体型小,成长速度慢,在头几年多以无脊椎动物、死亡的生物为食。这些特性可能是鳄鱼能够存活过白垩纪末灭绝事件的关键。
在灭绝事件过后,造成许多空缺的生态位,生态系统花了长时间才恢复原本的多样性。
微生物群
在钙质微型浮游生物的化石记录中,白垩纪末灭绝事件是最大规模的集体灭绝之一;钙质微型浮游生物形成的钙质沉积层,也是白垩纪的名称来源。这次灭绝事件对钙质微型浮游生物的影响,只有到种的程度。 根据统计分析,这段时间的海洋生物物种数量降低,多是因为迅速的灭亡,而非物种形成的减少。 由于微生物的化石记录多由囊胞所构成,而不是所有的甲藻门(Dinoflagellate)都具有囊胞,因此白垩纪-古近纪界线时期的甲藻类化石记录并不明确,造成相关研究的差异。 2004年的一个研究,则认为当时的甲藻类并没有发生明显变动。
最早自奥陶纪之后,放射虫(Radiolaria)一度消失于化石记录。在白垩纪/第三纪交界,可以找到少量的放射虫化石。没有证据显示当时的放射虫遭到大规模的灭绝。在古新世早期,放射虫在南极区相当繁盛,这可能因为寒冷的气候。 接近46%的硅藻物种,继续存活到上古新世。这显示白垩纪末的硅藻灭绝程度并不严重,层度仅到种的程度。
自从1930年代以来,科学家已着手研究白垩纪与第三纪交接时的有孔虫门(Foraminifera)灭绝状况。 自从撞击事件可能造成白垩纪末灭绝事件的理论出现后,更多科学家研究白垩纪-古近纪界线时的有孔虫门灭绝状况。 但科学界对此的意见分为两派,一派认为有孔虫门主要因为这次灭绝事件而影响, 另一派认为在白垩纪与第三纪交接时期,有孔虫门有过多次的灭绝与复原。
底栖的有孔虫类主要以生物有机碎屑为食,随者白垩纪末灭绝事件造成大量海洋生物死亡,众多的底栖有孔虫类也跟者灭亡。在海洋生态圈复原后,底栖有孔虫类的食物来源增加,物种形成也跟着增加。 浮游植物在古新世早期复原,提供大型的低栖有孔虫类食物。直到灭绝事件后的数十万年,约在古新世早期,低栖生态系统才完全复原。
海生无脊椎动物

一个菊石亚纲的化石
化石证据显示,在白垩纪﹣第三纪灭绝事件的前后,海生无脊椎动物有相当大的变化。但化石纪录的缺乏,使得无法显示出真实的灭绝比例。
介形纲(Ostracoda)是甲壳亚门的一纲,这群小型动物在马斯特里赫特阶上层相当繁盛,在多个地区留下化石纪录。一个针对介形纲化石的研究,显示介形纲在古新世时期的多样性,低于第三纪的其他时期。但目前的研究无法确定介形纲的衰退,是在白垩纪末事件的之前或之后发生。
在白垩纪晚期的珊瑚纲石珊瑚目中,有接近60%的属消失,没有存活到古新世。近一步的研究显示,栖息于温暖、副热带、浅海地区的珊瑚,记有98%的种灭亡。而那些栖息于较寒冷、透光带以下、单独生长、无法形成珊瑚礁的珊瑚,受到灭绝事件的影响较小。与进行光合作用的海藻共生共存的珊瑚,则在白垩纪﹣第三纪灭绝事件中大量灭亡。 但除了研究白垩纪-古近纪界线到古新世地层中的珊瑚化石以外,还要参考当时的珊瑚礁生态系统变化。
在白垩纪-古近纪界线之后,头足纲、棘皮动物、双壳纲的属数量,呈现明显的减少。 而大部分的腕足动物,在白垩纪末灭绝事件中存活,并在古新世早期开始多样化。

一个双壳纲中的厚壳蛤类化石,发现于阿拉伯联合酋长国的白垩纪末地层。图中的量尺长10毫米。
在软体动物头足纲中,大部分的物种在这次灭绝事件灭亡,只有鹦鹉螺亚纲(以现代的鹦鹉螺为代表)与鞘亚纲(演化成现代的章鱼、鱿鱼、乌贼)存活至今。这些灭亡的物种包含独特的箭石目,以及高度多样性、分布广泛且数量众多的菊石。研究人员指出鹦鹉螺亚纲产的卵较大、数量较少,是它们存活下来的重要原因。而菊石产的卵数量众多、以浮游生物方式生存,因此在白垩纪末灭绝事件中灭亡。在全球各地的菊石灭亡之后,鹦鹉螺类的外壳开始辐射演化,演化出不同的形状,构造日益复杂,与昔日的菊石类似。
在棘皮动物中,接近35%的属在这次灭绝事件中消失。其中,生存在低纬度、浅水的棘皮动物灭亡比例最高;而生存在中纬度、深海区域的棘皮动物,被影响的程度则较小。这种灭绝模式的成因可能是栖息地的消失,尤其是浅水地区的碳酸盐岩台地。
其他的无脊椎动物,例如厚壳蛤类(可形成礁的蛤蜊)与叠瓦蛤(现代扇贝的大型近亲),则在白垩纪﹣第三纪灭绝事件中全数灭亡。
鱼类
在白垩纪-古近纪界线的上下地层,有大量的有颌鱼类化石,可供科学家研究这些海生脊椎动物的灭绝模式。在软骨鱼纲中,有接近80%的鲨鱼、鳐目、?鱼的科存活过这次灭绝事件; 在真骨下纲中,只有少于10%的科消失。 在南极洲附近的西摩岛,化石纪录显示白垩纪-古近纪界线的上层有大量的硬骨鱼死亡。科学家推论当时的鱼类曾经面临环境变动,而K-T事件应是这次环境变动的主因。 但是,海洋与淡水环境,减轻了鱼类所遭受环境变动的影响。
陆地无脊椎动物
在北美洲的14个地点,开花植物化石被用来研究昆虫的多样性变化与灭绝比例。研究人员发现,在白垩纪-古近纪界线前的白垩纪地层,有丰富的植物,昆虫很多样化。在古新世早期(约灭绝事件后170万年),植物群仍相当多样化,来自昆虫的采食则较少。
陆地植物
在白垩纪﹣第三纪灭绝事件发生时,没有广布于全球的植物可供研究。 但个别地区的植物群连续状况可供研究。在北美洲,白垩纪﹣第三纪灭绝事件发生时有大量植物灭亡,但在该次灭绝事件发生前,已有明显的植物群变化。
在南半球的高纬度地区,例如新西兰与南极洲,植物群的物种并没有发生大量消失,但在植物的繁盛状况则有短期、剧烈的变化。 在北美洲,有接近57%的植物物种灭亡。在古新世,化石记录中的蕨叶,显示蕨类开始再度兴盛,植物群开始复原。在1980年的圣海伦火山爆发后,也出现类似的蕨类复兴状况。
由于白垩纪末灭绝事件,对植物群造成整体性的破坏,当时的腐生生物大量激增,例如真菌,这些生物不需依靠光合作用,而分解死亡植物以吸收营养。真菌的繁盛只持续了数年,当大气层恢复正常后,光合作用植物(例如蕨类)开始再度生长。
两栖动物
证据显示,两栖动物没有因为白垩纪﹣第三纪灭绝事件而大量灭亡,大多数成员都存活下来。 数个针对蒙大拿州的蝾螈化石研究指出,七个蝾螈属中有六个属存活过这次灭绝事件。
青蛙存活到古新世,似乎只有少数种灭亡。但在化石纪录中,青蛙的科与属数量并不清楚。 一个针对蒙大拿州的三个属的青蛙研究指出,它们并没有受到白垩纪末灭绝事件的影响。 这些数据显示,没有两栖类的科在这次灭绝事件中灭亡,或是受到打击。原因是两栖类可在水中生存,或者在沉积物、土壤、树木中筑穴、或是在岩层中的洞存活。
非主龙类的爬行动物
在白垩纪﹣第三纪灭绝事件中,非主龙类的爬行动物有龟鳖目、鳞龙超目(包含现今的蛇、蜥蜴、蚓蜥)存活下来,此外还有半水生的离龙目(属于原始的主龙形下纲,消失于中新世)也幸存过这次灭绝事件。 有超过80%的白垩纪乌龟,存活过这次事件。在白垩纪末,总计有六个乌龟的科,都存活到第三纪,并存活至现今。
现存的鳞龙类包含:喙头蜥目、有鳞目。在中生代早期,喙头蜥目是群相当成功的动物,而且分布广泛,但在白垩纪中期开始衰退。喙头蜥目目前仅存一属,仅存于新西兰。
现存的有鳞目包含:蜥蜴、蛇、以及蚓蜥,它们在侏罗纪占据了许多不同的生态位,并成功地存活到白垩纪。有鳞目存活过白垩纪末灭绝事件,成为现今最成功、最多样化的爬行动物,目前总计有超过6,000个种。目前未知有任何陆生有鳞目的科,在这次灭绝事件中消失,化石证据也显示它们的数量没有明显的变化。这些陆生有鳞目动物的体型小、适应性强的代谢率、以及可改变栖息环境的能力,是它们在白垩纪晚期/古新世早期幸存的原因。
非主龙类的海生爬行动物,包含沧龙类、蛇颈龙类,它们是白垩纪时的优势海生掠食动物,但在白垩纪末灭亡。
主龙类
主龙类包含两个主要演化支,镶嵌踝类主龙包含鳄鱼与其近亲,鸟颈类主龙包含翼龙目、恐龙、以及鸟类。
鳄形超目
在马斯特里赫特阶的化石纪录中,鳄形超目有10个科存在者,其中5科在白垩纪-古近纪界线后消失, 其余5科存活到古新世。这些幸存的鳄鱼大多栖息于淡水与陆地环境,唯有森林鳄科同时存活于淡水与海洋环境。由此可看出有50%的鳄鱼灭绝,而且没有大型鳄鱼存活下来,例如北美洲的恐鳄。 鳄鱼在灭绝事件后大量存活的原因,可能是它们的水生生态位与穴居的生活方式,这样可以减少环境剧烈变动的影响。 在2008年,Stéphane Jouve与其同事提出森林鳄科的幼年体生活于淡水环境,如同今日的海生幼年鳄鱼,所以当其他海生爬行动物灭亡时,森林鳄科仍得以继续幸存。淡水环境受到白垩纪﹣第三纪灭绝事件的影响,不若海洋环境来得大。
翼龙目
在马斯特里赫特阶,唯一确定存在的翼龙类是神龙翼龙科,神龙翼龙科也在灭绝事件中灭亡了。在白垩纪中期,仍有10科大型翼龙类存活者,而后逐渐衰退。较小型的翼龙类则早在马斯特里赫特阶以前就已灭亡。在马斯特里赫特阶前,已出现小型动物衰退,而大型动物更为繁盛的现象。在这个时期,新鸟亚纲开始多样化,并取代其他原始鸟类与翼龙类的生态位,这些动物之间可能出现直接的竞争,或只是新鸟亚纲占据这些消失物种的空白生态位。
恐龙

暴龙觅食的重建图
许多白垩纪末灭绝事件的研究,都关注于恐龙如何灭绝。除了少数的争议研究,大部分科学家同意恐龙(不包含鸟类)在白垩纪末灭绝事件中灭绝。关于恐龙的灭绝过程分为两派意见,第一派认为在白垩纪的最后数百万年,恐龙的多样性已出现衰退,第二派则认为,在最后数百万年,并没有迹象显示恐龙的衰退。而目前科学家仍无法根据这个时期的恐龙化石,在这两派意见中做出定论。 目前没有证据显示马斯特里赫特阶晚期的恐龙,有穴居、游泳、潜水等习性,所以无法减低环境变动所带来的影响。某些小型的恐龙可能会存活下来,但它们不分草食性或肉食性,都会面临植物或猎物的短缺。
近年来,关于恐龙是内温性动物的证据日渐增多,恐龙与近亲鳄鱼的不同代谢程度,有助于科学家研究它们在白垩纪末的灭绝与存活原因。鳄鱼属于冷血动物,可以连续数个月未进食;而恐龙体型接近的温血动物,需要更多的食物来源,以维持较快的代谢率。因此当K-T事件发生时,整个食物链崩溃,需要大量食物的恐龙灭亡,而鳄鱼继续存活。 鸟类与哺乳类等内温性动物,可能因为体型较小而所需食物较少,加上其他因素,得以在这次灭绝事件幸存。
有数个研究人员主张,恐龙的灭绝是逐渐性的,甚至有恐龙存活到古新世。他们宣称在海尔河组的岩层中,在白垩纪-古近纪界线的上方1.3米处,发现了7种恐龙牙齿化石,意味者在白垩纪-古近纪界线后的4万年,仍有恐龙存活者。 在犹他州圣胡安河的白杨山砂岩层(Ojo Alamo Sandstone),发现了鸭嘴龙类的包括股骨在内的34块骨骼化石,附近有古新世的标准花粉,也得到了古地磁证据的支持。科罗拉多州的阿尼马斯组(Animas Formation)含恐龙部分的时代也可能属于古新世。这个发现显示该种恐龙存活到第三纪,接近6450万年前,白垩纪-古近纪界线后的100万年。如果这个发现属实,这种鸭嘴龙类将成为幸免存活的演化支。 但是有些人研究认为,上述化石可能因为侵蚀作用,而被带离原本的地点,而在较年轻的沉积层中再度沉积。 2004年的一项研究表明在中国南雄盆地发现的鸭嘴龙化石和蛋化石中发现有第三纪的花粉沉积。
鸟类
大部分古生物学家认为鸟类是唯一存活到今日的恐龙。在鸟类中,除了新鸟亚纲以外的原始鸟类在白垩纪末灭绝,包含当时相当繁盛的反鸟亚纲与黄昏鸟目。 数个关于鸟类化石的研究显示,在白垩纪-古近纪界线以前,鸟类的物种已出现多样化现象,而鸭、鸡、以及平胸鸟类的近亲已经出现,并与恐龙共同生存。 许多新鸟亚纲因为能够潜水、游泳、或是在水中与沼泽寻找庇护地,因此得以在这次灭绝事件中存活。此外,许多鸟类能够筑穴,或生存在树洞或白蚁巢中,它们也存活过这次灭绝事件。这些存活下来的鸟类,在灭绝事件后占据许多恐龙的原本生态位。
哺乳动物

海尔河组
所有白垩纪的哺乳动物主要族系,包含:卵生的单孔目、多瘤齿兽目、有袋下纲、真兽下纲、磔齿兽超科(Dryolestoidea,原始的兽亚纲)、 以及冈瓦那兽亚目(Gondwanatheria,属于异兽亚纲), 都存活过白垩纪﹣第三纪灭绝事件,但都遭受重大损失。生存在北美洲与亚洲的原始有袋类三角齿兽超科(Deltatheroida)遭到绝种。 在北美洲的海尔河组,10种多瘤齿兽目中有至少一半成员,以及11种有袋类,都遭到灭亡。
在白垩纪﹣第三纪灭绝事件的前3000万年,哺乳动物开始多样化。白垩纪﹣第三纪灭绝事件阻碍了哺乳动物的进一步多样化。 目前的研究指出,尽管恐龙的消失留下大量生态位,但哺乳动物并未因此爆炸性多样化。 一个研究指出,数个哺乳动物的目,在白垩纪﹣第三纪灭绝事件后不久开始多样化,包含翼手目(蝙蝠)、鲸偶蹄目(鲸鱼与海豚、以及偶蹄类); 但另一个研究则提出不同看法,认为在白垩纪﹣第三纪灭绝事件后,只有有袋类开始多样化。
在K-T事件以前,哺乳动物的体型通常很小,接近老鼠的体型,使它们容易找到庇护地。此外,某些早期单孔目、有袋类、以及真兽类是半水生或穴居动物,如同许多现今哺乳动物的栖息环境。半水生或穴居哺乳动物可在白垩纪﹣第三纪灭绝事件造成的环境压力中找到庇护所。
证据
北美洲化石

在白垩纪﹣第三纪灭绝事件发生前的地层,有相当丰富的恐龙化石
在北美洲的陆相地层层序中,马斯特里赫特阶晚期的丰富孢粉型态与白垩纪-古近纪界线后的蕨叶,最能突显两个不同时期地层的差异。
在白垩纪-古近纪界线发生前,已知最富含恐龙化石的地层位于北美洲西部,尤其是美国蒙大拿州的海尔河组(年代为马斯特里赫特阶晚期)。若与海尔河组与加拿大亚伯达省与蒙大拿州的朱迪斯河组/恐龙公园组(年代较早,约7500万年前)相比较,可得出白垩纪最后1000万年的恐龙数目变化。这些地层的地理分布有限,仅分布于北美洲的部分地区。
在坎潘阶中到晚期的地层中,恐龙的多样性超过其他时期。马斯特里赫特阶晚期出现数种大型恐龙:暴龙、甲龙、厚头龙、三角龙、以及牛角龙, 显示在灭绝事件发生前,这个地区的食物相当丰富。
除了丰富的恐龙化石,植物化石纪录可重建出植物物种的大幅减少,成为灭绝事件的证据。在白垩纪-古近纪界线以上的地层,花粉化石证实该时期的优势植物是被子植物;在交界处的地层,花粉化石很少,主要是蕨叶化石。 从白垩纪-古近纪界线往上层探索,可以看出花粉化石的数量逐渐增多。在火山爆发后的地区,也可找到这种植物化石的改变模式。
海洋化石
海生浮游生物的大规模、迅速灭亡,发生于白垩纪-古近纪界线时。 早在白垩纪-古近纪界线前,菊石亚纲已有小规模、缓慢的衰退,可能与白垩纪晚期的海退有关,其余的属在白垩纪-古近纪界线时灭亡。双壳纲叠瓦蛤科的大部分物种,在白垩纪-古近纪界线前已经逐渐灭亡;同样在白垩纪末期,菊石的多样性也出现小规模、逐渐的衰退。 研究显示,在白垩纪晚期的海洋中,已出现数种不同的灭绝模式,发生时间部分交错,这些灭绝模式结束于白垩纪末灭绝事件。
经历时间
关于白垩纪﹣第三纪灭绝事件的持续时间,长久以来仍处于争议中,因为某些理论认为这次灭绝事件的过程很短(从数年到数千年之间),而其他的理论则认为灭绝的过程持续了长时间。由于不可能找到一个物种的完整化石纪录,所以已灭绝物种的真正灭绝时间,可能晚于最后的化石纪录。因此,灭绝事件的经历时间难以测定。 科学家已发现少数横跨白垩纪-古近纪界线、带有化石的地层,年代从白垩纪-古近纪界线前数百万年,到白垩纪-古近纪界线后近百万年。
灭绝原因
关于白垩纪﹣第三纪灭绝事件的成因,科学家们目前已提出数个理论。这些理论多关注于撞击事件,或是火山爆发,某些理论甚至认为两者都是原因 。在2004年,J. David Archibald与David E. Fastovsky试图提出一个结合多重原因的灭绝理论,包含:火山爆发、海退、以及撞击事件。在这个理论中,白垩纪晚期的海退事件,使陆地与海洋生物群落面临栖息地的改变或消失。恐龙是当时最大的脊椎动物,首先受到环境改变的冲击,多样性开始衰退。火山爆发喷出的悬浮粒子,使得全球气候逐渐冷却、干旱。最后,撞击事件导致依赖光合作用的食物链崩溃,并冲击已经衰退的陆地食物链与海洋食物链。多重原因理论与单一原因理论的差别在于,单一原因难以达成大规模的灭绝事件,也难以解释灭绝的模式。
撞击事件

小行星撞击地球的过程想像图
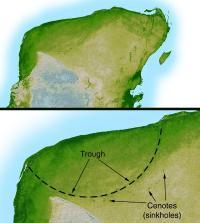
位于犹加敦半岛的希克苏鲁伯陨石坑,直径达180公里。
在1980年,诺贝尔物理学奖得主路易斯·阿尔瓦雷茨、与儿子沃尔特·阿尔瓦雷茨、弗兰克·阿萨罗(Frank Asaro)、海伦·米歇尔(Helen Michel)等人发现,全球的白垩纪与第三纪交接地层,地层中的铱含量高于正常标准;举原始研究中的两个地层为例,铱含量分别是正常标准的30倍、130倍。铱(Ir)是种地球地壳中非常少见的金属,属于亲铁元素;大部分的亲铁元素在行星分化的过程中沉降到地核部分,例如铁。由于大部分小行星与彗星常发现铱元素,阿尔瓦雷茨等人认为在白垩纪与第三纪的交接时期,曾有颗小行星撞击地球表面。 早在1950年代时,已经有类似的撞击事件理论,但当时没有发现任何证据。
在2007年9月,美国科罗拉多州美国西南研究院的研究人员威廉·波特克(William Bottke)与数位捷克籍的科学家,利用电脑计算出希克苏鲁伯撞击事件的可能肇事者。根据他们的模拟计算结果,该陨石坑有90%的可能性是由一个名为巴普提斯蒂娜的小行星所造成的,该小行星直径接近160公里,目前运行于火星与木星之间;根据推论,在1亿6000万年前,巴普提斯蒂娜小行星被一颗直径约55公里的未命名小行星撞击后粉碎,主要的碎片形成巴普提斯蒂娜族小行星带,而某些碎片则闯入地球的公转轨道,其中一颗直径10公里的碎片,在6,500万年前撞击了墨西哥尤卡坦半岛,形成希克苏鲁伯陨石坑。另外,月球表面的第谷环形山形成于1亿800万年前,也可能是巴普提斯蒂娜族小行星带的撞击结果。
撞击事件会造成大量的灰尘进入大气层,遮辟阳光,时间长达1年或1年之内。另外,大量的硫酸以气溶胶的形式进入平流层中,使抵达地表的日照量降低10%到20%,妨碍植物进行光合作用;平流层内的气溶胶完全沉降,可能需要10年之内的时间,这将导致植物、浮游植物的大量消失,而在食物链上层的草食性动物、掠食性动物也跟着灭亡。而以生物有机碎屑为食的小型生物,受到灭绝事件的影响较小。 撞击事件后的数小时内,会有大量的红外线穿透大气层,对曝露在外的生物造成伤害。 撞击与再度落下的喷出物,会造成全球性的火风暴。白垩纪晚期的大气层氧含量高,会助长各地的火势;而第三纪早期的大气层氧含量,呈现下降的迹象。如果当时曾发生全球性的火风暴,大气层中的二氧化碳含量将上升,并在空气中的灰尘、微粒沉降后,继续造成短期的温室效应。在撞击事件时存活的生物,可能因上述效应而死亡。
撞击事件可能会造成酸雨,这依撞击发生地点的地层成分而定。但科学家指出,酸雨造成的影响相对而言较小,而且最多持续约12年。 大自然环境会将酸雨稀释、中和,而且灭绝事件的部分幸存者对酸雨相当敏感,例如青蛙,显示酸雨并非灭绝事件的重要因素之一。撞击产生的灰尘、可能还有带硫的气溶胶,会在大气层中慢慢沉降,最多不超过10年,所以只能解释迅速的灭绝原因。 位于墨西哥尤卡坦半岛沿岸的希克苏鲁伯陨石坑,是最符合阿尔瓦雷茨理论的陨石坑。在1978年,格伦·彭菲尔德(Glen Penfield)发现并进行希克苏鲁伯陨石坑的鉴定工作。在1990年,其他研究人员完成希克苏鲁伯陨石坑的研究,该陨石坑呈椭圆形,平均直径为180公里,规模符合阿尔瓦雷茨理论的推测。
从陨石坑的地点与形状判断,该小行星撞击到陆地与海洋交界,除了造成大量灰尘与微粒,还有大型的海啸。加勒比海与美国东部的数个地层,在当时为陆相地层,却发现来自海洋的砂;在其他的海相地层,则发现来自陆地的植物碎屑与岩石。撞击发生处有碳酸钙地层,这将产生大量的二氧化硫气溶胶,也会降低地表的日照量,造成酸雨,对植物、浮游生物、具有钙质外壳的生物造成伤害,例如颗石藻与软体动物。在2008年8月,一群科学家利用地震学资料,提出该陨石撞击到的水域,比往常所认定的更深。他们提出这将造成更多的硫酸气溶胶,严重影响气候,造成更多酸雨。 大部分的古动物学家同意在6,500万年前曾有发生小行星撞击地球,但撞击事件是否还有其他因素,仍在研讨、探究中。
德干暗色岩
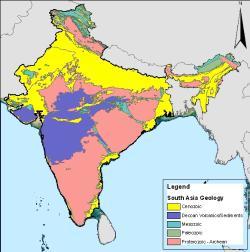
印度的岩石年代图,粉红色部分为德干暗色岩爆发形成的火成岩。
在2000年以前,就有德干暗色岩洪流玄武岩是这次灭绝事件的主因,但因为这次火山爆发始于6,800万年前,并持续超过200万年,所以当时认为德干暗色岩火山爆发所造成的灭绝事件应该是逐步、缓慢的。但现在有证据显示,有2/3的德干暗色岩地形,是在一万年内形成的,发生于6,550万年前。显示这次火山爆发造成了快速的灭绝事件,可能在数千年内,但还是远比撞击事件长 。
德干暗色岩火山爆发借由数种方式造成了灭绝事件,灰尘与二氧化硫与二氧化碳等温室气体大量喷出,灰尘使地表的日照量下降,植物的光合作用减少;当灰尘慢慢降落至地表时,火山爆发喷出的二氧化碳气体,可能增加了温室效应。许多植物因此而死亡,草食性恐龙因没有食物而灭亡,而肉食性恐龙也相继灭绝 。在恐龙大规模灭绝前,火山爆发喷出的气体,似乎与全球暖化有关。有证据指出在希克苏鲁伯撞击事件之前的50万年内,大气温度曾上升8 °C 。
在德干暗色岩火山爆发仍被认为是缓慢灭绝事件时,路易斯·阿尔瓦雷茨便已提出古生物学家被少数的资料所混淆。他的警告当时被没有被接受,但后来针对化石层的研究工作,证明早期的相关研究有误。目前大部分古生物学家已普遍接受撞击事件是白垩纪末灭绝事件的主要原因。不过,路易斯·阿尔瓦雷茨的儿子沃尔特·阿尔瓦雷茨(Walter Alvarez)认为在这次撞击事件前,地球上已发生其他重大事件,例如:海平面下降、造成德干暗色岩的大规模火山爆发,它们可能也是这次灭绝事件的综合原因之一 。
多重撞击事件
目前已经发现数个陨石坑,形成时间相当于白垩纪-古近纪界线。这些发现指出,当时可能有多次撞击事件发生;这种模式类似发生在1994年的苏梅克-列维9号彗星,因为潮汐力而分裂,数个碎片在接下来的几天内陆续撞击木星。这些陨石坑包含:乌克兰的波泰士陨石坑,直径24公里,形成时间约在距今6517万年前(误差值为64万年)、北海的银坑陨石坑,直径20公里,形成时间约在距今6,500万年前到6,000万年前。位于印度外海的湿婆陨石坑,当时位于特提斯洋,也是多重撞击事件的可能陨石坑之一。
马斯特里赫特阶的海退事件
有明确的证据显示,在白垩纪马斯特里赫特阶发生大规模的海退,达到中生代以来最低的程度。在世界各地的一些马斯特里赫特阶地层,最早的部分是海床,较早的部分是海岸地层,最晚部分则是陆相地层。这些地层没有因为造山运动而倾斜、折曲的迹象,因此最有可能的解释是海退。目前没有海退原因的相关证据,较为普遍的解释是中洋脊的活动降低,而这些巨大海底山脉随者自身的重量而缓慢沉降于海底地层中。
大规模海退造成大陆架大幅消失,栖息在大陆架(或称陆棚、大陆架)的海洋生物最丰富,因此海退可能会造成海洋生物的灭绝。研究也显示,海平面的变化,不足以造成如此严重的菊石灭亡。海平面的下降可能造成大气环流系统与洋流系统的变化,形成气候变迁;海洋面积的缩小也会使地表的反照率下降,而使全球气温上升。
大规模海退也使许多大陆海消失,例如北美洲的西部内陆海道。这些海域的消失,破坏许多存在于1000万年之前、生物繁盛的海岸平原,例如恐龙公园组。同时,由于陆地相对上升,河流的长度更长,使淡水的生存区域扩张。海洋与淡水区域的消长变化,使淡水脊椎动物增加,而海洋生物则数量减少,例如鲨鱼。
参考资料
(英文) Fortey R. Earth: An Intimate History. Vintage. 2005. ISBN 0375706202.
(英文) BBC新闻:恐龙的灭绝原因是?科学家怀疑除了陨石撞击以外,还有其他的元凶。
(英文) 国家地理频道:杀死恐龙的元凶共有四个:气候变化、火山爆发、希克苏鲁伯陨石坑、以及另一个年代较晚的不明撞击事件。
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

相关资料





- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读





关于我们

APP下载


