





-
 QQ空间
QQ空间
-
 QQ好友
QQ好友
-
 微信好友
微信好友
-
 新浪微博
新浪微博
主龙类
译名
主龙类(Archosauria)在希腊文意为“具优势的蜥蜴”。但许多中文网站与书籍,误认学名的意思为“古老的蜥蜴”,而翻译为初龙类、祖龙类、古龙类。
在某些中文网站中,仍然保持早期的传统分类法,将主龙类(Archosauria)分类成双孔亚纲的一个下纲,名为主龙下纲、初龙下纲;或是爬行纲的一个亚纲,名为主龙亚纲、初龙亚纲、祖龙亚纲、古龙亚纲。
特征
主龙类的最基本且最广为接受的共有衍征包括:
牙齿位在齿槽内,这让它们进食时牙齿较不易脱落(这也是槽齿目的名称来源。不过目前槽齿目被视为早期主龙类的并系群集合,不被现代古生物学家所接受)。
眶前孔(Preorbital fenestrae,位于鼻孔与眼睛之间)减轻了头颅骨的重量,因为大部分早期主龙类的头颅骨长而重,这对主龙类是一个非常重要的特征。眶前孔通常大于眼眶。
下颌孔(Mandibular fenestrae,下颌骨头上的小洞孔)稍为减轻了下颌的重量。
股骨有第四粗隆部(肌肉附着点)。这个特征似乎并不重要,却是恐龙演化出直立姿态的关键之一(所有早期恐龙以及许多晚期恐龙是二足动物),也可能与主龙类或它们的祖先在二叠纪-三叠纪灭绝事件中存活下来有关联。
主龙类在三叠纪兴起

鸟颈类主龙的西里龙、镶嵌踝类主龙的波罗尼鳄
在二叠纪时期,似哺乳爬行动物是陆地上的优势脊椎动物,但大部分在二叠纪-三叠纪灭绝事件中灭亡。草食性的似哺乳爬行动物水龙兽是唯一存活下来的大型陆地动物,并在三叠纪初期成为最繁盛的陆地动物 。
在早三叠纪,主龙类快速地成为陆地上的优势脊椎动物。关于主龙类为何快速崛起,最常提出的两个解释分别为:
主龙类演化出直立四肢的过程,比似哺乳爬行动物的演化还快;这让它们有更多的耐力,可以在运动时同时呼吸。但这论点是有问题的,因为主龙类拥有优势地位时,它们仍处于躺卧或半直立姿势,类似水龙兽或其他似哺乳爬行动物。
早三叠纪的气候环境普遍干燥,这是因为当时的陆地集合为盘古大陆。主龙类的保存水分能力,比似哺乳爬行动物好:
现代双孔动物(蜥蜴、蛇、鳄鱼、鸟类)都将尿酸以糊状排泄出来。根据合理的推测,主龙类也将尿酸排泄出来,因此能更好地保存水分。而双孔动物缺乏皮腺的皮肤也有助于保存水分。
现代哺乳类排泄尿素,需要大量的水分以溶解尿素。皮肤也布有许多皮腺,会流失许多水分。根据推测,似哺乳爬行动物的皮肤拥有类似的特征,但这仍在争论中,详见Palaeos网站。因此它们在干燥的环境中拥有劣势。在澳洲的一个地层显示出澳洲的上新世至更新世期间,气候可能一样干燥,而该地最大型的陆地掠食动物是古巨蜥(Megalania)与陆地鳄鱼,而非哺乳类。
主要的种类
自从70年代以来,科学家主要依据主龙类的踝部结构来将它们分类。
最早的主龙类拥有原始的中跗骨踝部:距骨与跟骨借由关节与缝合处固定至胫骨与腓骨。
镶嵌踝类主龙出现于早三叠纪。它们的距骨借由关节与缝合处与胫骨连接,而距骨沿者关节嵌入跟骨的窝槽。早期镶嵌踝类主龙仍然以采取四肢往两侧伸展的步态,但有些晚期镶嵌踝类主龙发展出完全直立的四肢(尤其是劳氏鳄目)。现代鳄鱼也属于镶嵌踝类主龙,可依据它们的行走速度,来采用四肢往两侧伸展或四肢直立的步态行走。
派克鳄以及鸟鳄科拥有与镶嵌踝类主龙不同的反转型态踝部,而距骨上有窝槽,而跟骨上有骨桩。
最早的鸟颈类主龙化石发现于晚三叠纪卡尼阶,但目前不清楚鸟颈类主龙如何从镶嵌踝类主龙演化而来,它们也可能较早演化出现,也可能从具有原始中跗骨的早期主龙类演化而来。鸟颈类主龙的先进中跗骨形态,是由非常大的距骨与非常小的跟骨所构成,可使踝部关节在单一平面上移动,如同简易的铰链。这种结构仅适合拥有直立四肢的动物,并在它们奔跑时提供更多稳定性。鸟颈类主龙与其他主龙类的差别在于:它们体型较轻巧,通常为小型、它们的颈部较长,并呈S形弯曲、它们的头颅较为轻巧、许多鸟颈类主龙是完全二足动物。鸟颈类主龙与其他主龙类的差别在于:身体通常较小、体格轻、颈部长而呈S形弯曲、头颅骨较为轻型、而且大部分为二足动物。主龙类的股骨具有第四粗隆部,可使鸟颈类主龙更易演化成二足步态,并提供大腿肌肉更多的力量。在晚三叠纪期间,鸟颈类主龙进一步分化成翼龙目与恐龙。
臀部关节与运动方式
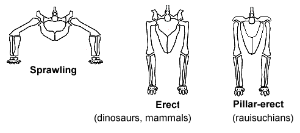
三种不同的臀窝结构与后肢直立方式,从左到右分别为往两侧延展、直立(哺乳类与恐龙)、柱状直立方式(劳氏鳄目)
如同早期四足总纲动物,早期主龙类的四肢姿势为往两侧延展。原因为:
它们的臀窝(Hip sockets)朝向两侧。
股骨顶端的头与股骨本身排列为一直线。
在三叠纪早期到中期,有些主龙类演化支独自发展出可直立的臀部关节。这个特征使它们拥有更多的活动力,而并避免在快速移动时无法同时呼吸。主龙类发展出两种型态的可直立关节:
臀窝朝向两侧,与股骨头接合。恐龙发展出这种直立方式。
臀窝朝向下方,股骨头与股骨本身平行。不同的主龙类演化支似乎多次个别演化出这种“柱状直立方式”,例如劳氏鳄目与某些坚蜥目。
灭绝与兴盛
鳄目、翼龙目、恐龙、以及离龙目在1亿9500万年前的三叠纪-侏罗纪灭绝事件中存活下来,但其他早期的主龙类则灭绝了,例如植龙目与劳氏鳄目。
恐龙与翼龙目在白垩纪-第三纪灭绝事件中灭亡,但鳄目、离龙目、以及鸟类存活下来。鸟类是主龙类的后代,因此在种系发生学中,鸟类属于主龙类。离龙目则在渐新世时期灭亡。目前只有鳄目(包括现代鳄鱼、长吻鳄、短吻鳄)与鸟类繁盛于现代。
主龙类的生活方式
食性
大部分主龙类是大型掠食动物,但有不同演化支演化成其他的生态位。坚蜥目是草食性动物,有些发展出鳞甲。少数鳄目是草食性动物,例如狮鼻鳄(Simosuchus)、叶齿鳄。早白垩纪的大型腔鳄可能是滤食性动物。蜥脚形亚目、鸟臀目是草食性动物,演化出多样性的摄食特征。
栖息地
主龙类主要为陆栖动物,以下例外:
鳄目是河流与沼泽里的优势动物,甚至进入到海中(真蜥鳄科、地蜥鳄科、森林鳄科)。地蜥鳄科甚至具有类似海豚的外形、桨状的前肢、锚状的尾巴、以及平滑无装甲的皮肤。
翼龙目与鸟纲演化出飞行能力,是优势飞行动物。
代谢
主龙类的代谢仍处于争议中。它们的确从冷血动物演化而来,而目前存活的非恐龙主龙类,鳄鱼,是冷血动物。但鳄鱼拥有一些与温血动物相关的特征,因为它们的促进氧气供给方式:
2个心室与2个心房。哺乳类与鸟类也有2个心室与2个心房。非鳄鱼的爬行动物的心脏有3个心房心室,这样比较没有效率,因为这样会将含氧血液与缺氧血液混合在一起,因此会将部分缺氧血液送至身体各处,而非送到肺脏。现代鳄鱼虽然拥有四腔室心脏,但与身体相比比例较小,并且与现代哺乳类与鸟类相比,血压较低。它们也拥有分导管,可让它们位在水面下时,以三腔室心脏运作,以储存氧气。
次生颚,可允许动物在进食时可以同时呼吸。
肝瓣,是肺脏的呼吸推动装置。与哺乳类与鸟类的肺脏推动装置不同,但根据某些研究人员宣称,较为类似某些恐龙。
有些专家认为鳄鱼最初为活耀、温血的掠食动物,而它们的主龙类祖先也是温血动物。研究显示鳄鱼的胚胎具有四腔室心脏,成长后改变为三腔室心脏,以适应水中环境。这些研究人员根据胚胎重演律,它们提出最初的鳄鱼具有四腔室心脏,因此它们为温血动物,而后来的鳄鱼发展出旁管,重新成为冷血动物,以及水底中的伏击掠食动物。
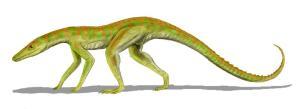
陆鳄是种非常小型、行动敏捷的鳄形类,生存于三叠纪晚期。
如果最初的鳄鱼、以及其他的镶嵌踝类主龙都是温血动物,这样可以解决一些演化的谜题:
最早期的鳄类,例如陆鳄,是种纤细、长腿的陆地掠食动物,生活方式可能相当活跃,这样需要非常快速的新陈代谢。而其他的镶嵌踝类主龙似乎拥有直立的四肢,例如劳氏鳄目。直立的四肢有益于活跃的动物,可让它们快速运动时,可以同时呼吸;但不利于缓慢的动物,因为身体站立或趴下时将消耗能量。
如果早期主龙类是完全冷血动物,恐龙将以短于似哺乳爬行动物的一半时间,演化为为温血动物。
呼吸系统
在2010年的一项研究,发现美国短吻鳄有独特呼吸方式。这项研究发现,当美国短吻鳄吸气时,的空气会进入第二支气管,流经交换气体的第三支气管,最后进入第一支气管。当美国短吻鳄呼气时,空气会经由第二支气管、第三支气管、第一支气管,最后从气管呼出体内。现代鸟类也有类似的呼吸方式,在呼吸过程中,空气以单向方式在气囊内流通;而哺乳动物的呼吸方式,则是反复方向的空气流通,气体在肺泡交换。除了现代鸟类以外,许多恐龙演化支也被发现拥有气囊,可以增进它们的呼吸效能。
由于现代鳄鱼、鸟类都有类似方式的呼吸方式、辅助呼吸系统,有可能是三叠纪的原始主龙类已经演化出这种呼吸系统,而大部分后代都有类似呼吸系统,例如:恐龙、翼龙类、鳄形超目…。研究人员也推测,主龙类的特殊呼吸方式,也是它们与似哺乳爬行动物的兢争过程中,逐渐占优势的原因之一。在三叠纪时期,大气层中的含氧量较低,主龙类的特殊呼吸方式,使呼吸效能较高,较具竞争优势。
种系发生学
简易版主龍類 Archosauria |--鑲嵌踝類主龍Crurotarsi | |-?梳棘龍科Ctenosauriscidae | `--鱷型踝關節類Crocodylotarsi | |--鳥鱷科Ornithosuchidae | `--+--植龍目Phytosauria | `--Suchia | |--迅猛鱷科Prestosuchidae | `--Rauisuchiformes | |--堅蜥目Aetosauria | `--勞氏鱷目Rauisuchia | |--勞氏鱷科Rauisuchidae | `--+--Paracrocodylomorpha | `--鱷形超目Crocodylomorpha (鱷魚與其近親) `--鳥頸類主龍Ornithodira |--Pterosauromorpha | |--Scleromochlus | `--翼龍目Pterosauria `--恐龍形態類Dinosauromorpha `--恐龍形類Dinosauriformes `--恐龍總目Dinosauria |--鳥臀目Ornithischia `--蜥臀目Saurischia `--鳥綱Aves
参考资料
Michael J. Benton (2004), Vertebrate Paleontology, 3rd ed. Blackwell Science Ltd
Robert L. Carroll (1988), Vertebrate Paleontology and Evolution, W. H. Freeman and Co. New York
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。




- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读

知识互答

关于我们

APP下载



{{item.time}} {{item.replyListShow ? '收起' : '展开'}}评论 {{curReplyId == item.id ? '取消回复' : '回复'}}