





-
 QQ空间
QQ空间
-
 QQ好友
QQ好友
-
 微信好友
微信好友
-
 新浪微博
新浪微博
真菌
特征
在引进用系统发生分析分子的方法之前,分类学家认为真菌是因为在生活方式相似的植物界的成员:真菌和植物两者都是不能移动的,并且在一般的形态和生长的栖息地的相似之处。像植物一样,真菌常常生长在土壤,并且在蘑菇的情况下,形成明显的子实体,有时像植物,例如苔藓。现在的真菌界被视为一个独立于植物界和动物界之外的生物界,大约从十亿年前就有显着遗传分化 。
形态学
微观的结构
大多数真菌成长为菌丝,这是圆柱形的线状结构,直径为2-10微米,和长度可达数厘米。菌丝生长在它们的顶端(顶点);新菌丝通常通过沿现有菌丝新梢被形成出现,被称为分支过程,或偶尔生长菌丝分叉,从而产生了两个平行的生长的菌丝 。
宏观的结构
真菌菌丝体可以成为可见肉眼的,例如,各种表面和基质,如墙壁和变质食品,其中,它们是通常被称为霉菌。 蕈类包括菌柄、菌盖和产生孢子的菌褶等三个部分。菌盖高于地面,这样散落的孢子就可以被空气气流带走。菌柄的底部有许多叫作菌丝的细线,蕈类就是通过菌丝来寻找养料的。
生殖
真菌的繁殖是复杂的,反映了生物的这个多样的生物界的生活方式和基因构成的差异 。据估计,三分之一的全部真菌的使用一种以上的方法传播;例如,可以在一个物种生命周期内发生的两个分化良好的阶段,可能是有性型和是无性型的生殖 。环境条件触发基因决定的,导致创立有性的或无性的繁殖的专用结构的发展状态。
真菌的生殖有有性生殖和无性生殖时期。
分类
真菌过去通常被视为植物的一种,因其型态和生境皆和植物相似:通常生长在泥土、不会移动;自1969年五界系统出现后,真菌已成为独立的“界”,与植物、动物和原生生物并列。
透过分子亲缘关系学的分析,真菌和动物一样为后鞭毛生物内的单系群 ,亦即真菌和动物的共祖是 单鞭毛生物 的后代——后鞭毛生物,而植物的祖先则是 双鞭毛生物 ,所以真菌和动物的关系比和植物的更亲近。真菌的分类是不断变动的,特别是在以DNA比对为基础的最近研究之中。最近在亲缘关系学上的研究时常会推翻之前由形态特征及由实验配种获得的物种概念等旧方法得来的分类。
在高层的分类单元中还不存在一个唯一且被广泛接受的系统,并且在每一层之中都会有频繁的名称异动,由物种上至门。学者们现在正在努力建立且促使一个统一且更一致的命名法的使用 。真菌的物种依其生命周期及生殖方法(有性或无性),可以有两种以上的科学名称。一些如真菌索引及ITIS之类的网站有列出真菌物种的最新名称(交叉参酌旧式的命名)。
2007年真菌界分类法是许多真菌学家及其他研究真菌分类学的科学家们进行大规模合作研究所得来的成果 。此分类法将真菌界分成七个门,其中的子囊菌门及担子菌门归于双核亚界之中。下边的分支图描绘出真菌界主要的分类单元,以及真菌界和后鞭毛生物与单鞭毛生物之间的关系。分支的长度并不正比于演化的长度。
20世纪的七、八十年代,黏菌(现属于变形虫界或归于原生生物界)、卵菌及丝壶菌(被归于囊泡藻界不等鞭毛门或原生生物界)不再列入真菌的范围。部分真菌学(mycology)与真菌学家(mycologist)的研究范围仍包括上述物种。因此,现今把传统的广义真菌称为 菌物 。 中国植物学会真菌学会于1993年5月改组为中国菌物学会。
系统发生学
分类类群
真菌主要的门是由其生殖结构上的特征来分类的。现在,共分成七个门被提议::微孢子虫门、壶菌门、芽枝霉门、新美鞭菌门、球囊菌门、子囊菌门及担子菌门 。

显微镜下的丛枝菌根,长在亚麻根部的皮质细胞上。
亲缘关系学上的研究已证实微孢子虫门(动物及单细胞生物上的单细胞寄生虫)是相当近期且高度衍生的组织内寄生真菌(生存于其他物种的组织内) 。2006年的一项研究断定微孢子虫门为其他真菌的旁系群 。 Hibbett和其同事认为此一研究并不会和他们对真菌的分类产生冲突,虽然微孢子虫门提升到门这一分类单元,为了弄清楚此类群之内的演化关系,还需要更多的研究 。
壶菌门的真菌分布在世界各地。壶菌会产生游动孢子,可以在液体中以单一个鞭毛自主地移动,这使早期的分类学家误以为它们是单细胞生物。由核糖体内的rRNA序列做出的分子亲缘关系学研究推定,壶菌为其他(由四个推定为并系群,或可能为多系群的分支所组成的)真菌门的基本分支 。
芽枝霉门之前被认为是壶菌门内的一分支。然而,最近的分子资料及超微构造上的特征将芽枝霉门归成接合菌门、球囊菌门及双核亚界(子囊菌门及担子菌门)的旁系群。芽枝霉门是腐生生物,分解有机物质来获取养分,且为所有真核生物的寄生虫。不像其最亲近的壶菌门是行合子减数分裂,芽枝霉门行孢子减数分裂 。
新美鞭菌门之前归于壶菌门内。此门内的真菌为厌氧生物,生存于大型草食性哺乳动物的消化系统内,亦可能生存于其他的地上及水中环境里。新美鞭菌门缺乏线粒体,但含有氢化酶体。做为壶菌门的近亲,新美鞭菌门亦会产生游动孢子,其孢子有单一后鞭毛或多鞭毛等类型 。
球囊菌门会形成丛生菌根,会侵入植物的根部细胞,和植物共生,以增加养份的供给来使双方都获得好处。所有现知的球囊菌门的物种皆行无性生殖 。球囊菌门和植物之间的共生关系非常久远,可追溯至4亿年前之久 。球囊菌门之前为接合菌门的一部分,但在2001年提升为门,且在现在取代了接合菌门 。之前归于接合菌门的真菌现在被重新归于球囊菌门,或地位未知的亚门(毛霉菌亚门、梳霉亚门、捕虫霉菌亚门及虫霉菌亚门)之中 。和医学相关的属有毛霉属、根粘霉属及根霉属。
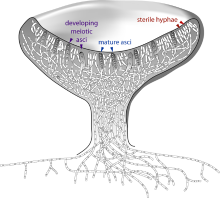
子囊果的图示
子囊菌门是真菌界中最大的分类类群 。此类真菌会产生减数分裂的孢子,称为囊孢子,被包在称为子囊的特化囊状构造中。子囊菌门包含牛肚菌、一些菇类及松露、单细胞的酵母(如酵母属、克鲁维酵母属、毕赤酵母属及假丝酵母属),以及其他生为腐生生物、寄生虫及共生生物的丝状真菌。丝状的子囊菌门生物中较显著及重要的属包括曲霉属、青霉菌、镰孢菌属及麦角菌属。许多子囊菌门的物种只被观察到行无性生殖,但在分子资料上的分析则时常能确认出其有性世代 。因为减数分裂的产品会留在囊状的子囊内,子囊菌门曾被用来说明遗传学的原理(如粉色面包霉菌) 。
担子菌门会在称为担子的柄上产生称为担孢子的孢子。最常见的菇类属于此类群,柄銹菌及会黑穗菌等主要的谷物病原体也一样属于此类群。其他重要的担子菌门生物亦包括玉米病原体(玉米黑穗菌) 、与人类偏利共生的属(马拉色菌属) 及会机会染人类的病原体(新型隐球菌) 。
类似真菌生物
因为在形态和生活方式的相似之处,粘菌(黏菌(mycetozoans),plasmodiophorids,acrasids,Fonticula和labyrinthulids,现在已经分别分类为变形虫门,有孔虫界,古虫界,后鞭毛生物和不等鞭毛类),水霉菌(卵菌)和hyphochytrids(均为原生藻菌)以前是的归类于真菌界,像Mastigomycotina,Gymnomycota和藻状菌纲组。粘菌也被作为原生动物研究,导致重复分类。
养分
真菌不能自行制造养分,因而无法独自于自然界生存,必须与别种生物借由寄生、腐生或者共生方式来吸取生活需要的营养。 它主要依靠营养菌丝(酵母菌除外)来吸收营养物质。真菌的菌丝有四种:气生菌丝、营养菌丝、匍伏菌丝、直立菌丝。
人类利用
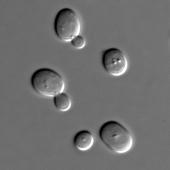
DIC显微镜 ( 英语 : Differential interference contrast microscopy ) 显示酿酒酵母细胞
人类利用菌类配制或保存食物等与其他用途是非常广泛的,并有着悠久的历史。蘑菇种植和蘑菇采集在许多国家是大型产业。
治疗用途
现代化疗
许多物种产生的代谢物是药理活性药物的主要来源。特别重要的是抗生素,包括青霉素,这是一种从小肽合成的β内酰胺类抗生素结构相关基团。尽管天然存在的青霉素如青霉素G(由产黄青霉菌生产)具有相对窄谱的生物活性,范围广泛的其他青霉素可由天然青霉素的化学改性来制备。现代青霉素是半合成的化合物,从最初发酵培养获得,但随后结构被改变为特定的期望性质 。
世界上第一种抗生素青霉素是青霉菌的分泌物。
传统和民间医药

药用真菌赤芝(左)和冬虫夏草(右)
某些蘑菇享有作为传统医学药物的使用,如中国传统医学药物疗法。带有应用的详细记录历史的值得注意的药用蘑菇包括姬松茸 ,灵芝 ,和冬虫夏草 。
发酵食品
面包酵母或酿酒酵母,一种单细胞真菌,是用来做面包等小麦制品,如比萨面团和饺子 。酵母属的酵母物种也可用于通过发酵生产酒精饮料 。
真菌修复
某些真菌,尤其是“白腐”菌,能降解杀虫剂,除草剂,五氯酚,杂酚油,煤焦油和重油并将其转化为二氧化碳,水和基本元素 。
其他
真菌被广泛用于生产工业化学品,像柠檬酸,葡萄糖酸,乳酸,苹果酸, 和工业酶,如用于 生物洗涤剂 ( 英语 : Biological detergent ) 的脂酶 ,用于制备纤维素乙醇 和 磨砂 ( 英语 : Stone washing ) 牛仔裤的纤维素酶和淀粉酶,蔗糖酶,蛋白酶和木聚糖酶。
参见
真菌分类表
真菌病
MycoBank
注释
^ 此分类系统是基于2007年Hibbett等人对亲缘关系学上的研究。
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。



- 有价值
- 一般般
- 没价值








24小时热门







推荐阅读


知识互答

关于我们

APP下载



{{item.time}} {{item.replyListShow ? '收起' : '展开'}}评论 {{curReplyId == item.id ? '取消回复' : '回复'}}