






古菌
历史
古菌知识图谱
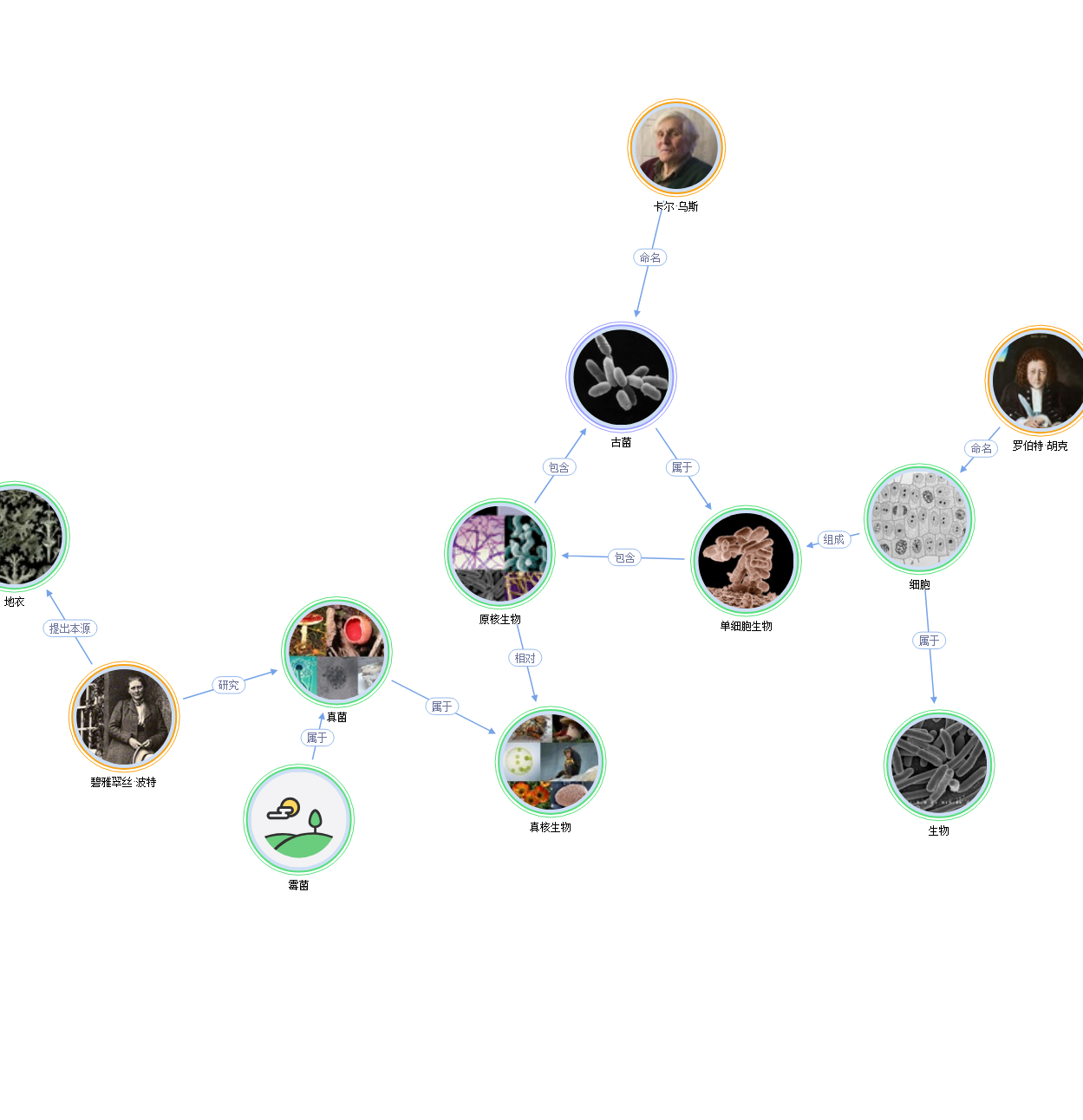
查看详细关系图古菌关系图谱
新域的诞生
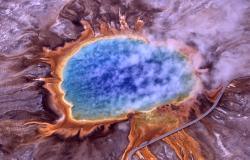
古菌最早是在一些极端环境,如火山温泉中发现的。图为黄石国家公园的大棱镜温泉。
在20世纪的大部分时间里,原核生物因其生化、形态和代谢上的一致性被视为分类上的单组生物。在这一时期,微生物学家试图根据其形状、细胞壁结构以及所消耗的物质来对微生物进行分类 。然而,在1965年有一种新方法被提出 ,即使用生物体的基因序列来搞清这些生物是如何相关联的。这种方法称为系统发育,是今天生物分类所使用的主要方法。

古菌给一些温泉(例如牵牛花温泉)染了色。
“古细菌”这个概念第一次出现是在1977年,由卡尔·乌斯和乔治·福克斯提出的,原因是它们在16S 核糖体RNA的系统发生树上和其它原核生物的区别 。这两组原核生物起初被定为古细菌和真细菌两个界或亚界(当时乌斯与福克斯更倾向于用“总界”一词)。后来乌斯认为它们是两支根本不同的生物,于是在1990年重新命名其为古菌和细菌 ,并将这两支和真核生物划为域这一新建立的分类阶元,一起构成了生物的三域系统。
起初只有一些 产甲烷菌 ( 英语 : methanogen ) 属于这个新域,这些古菌以一些极端环境为栖息地,比如温泉和盐湖。到20世纪末,微生物学家意识到古菌是一个庞大而多元化的群体,不只局限于极端环境,而是广泛存在于自然界中,如土壤和海洋里 。由于聚合酶链式反应(PCR)被大量用于探测采样土壤或水中的原核生物的核酸,古菌的重要性和独特性得到新的重视。PCR方法可以不经 细菌培养 ( 英语 : microbiological culture ) 而直接探测和鉴别微生物。
现行分类

ARMAN ( 英语 : archaeal Richmond Mine Acidophilic Nanoorganisms ) :2006年在西班牙力拓河的 矿渣酸性废水 ( 英语 : acid mine drainage ) 中发现的古菌新类群
古菌和原核生物的分类一般是一个快速发展并充满争议的领域。现行的分类体系旨在将古菌组织成具有共同结构特征和共同祖先的生物群 。这些分类很大程度上依赖于使用核糖体RNA基因序列来揭示生物之间的关系(分子系统发生学) 。目前大部分可纯化培养的并研究清楚的古菌都可以划为两个主要的门:泉古菌门和广古菌门。其它的门类还在初步创立阶段,比如2002年由Karl Stetter发现的奇特的物种 骑行纳古菌 ( 英语 : Nanoarchaeum equitans ) 而建立的纳古菌门 。还有1994年Barns等建议建立的一个新门初古菌门包括一小群不寻常的嗜热品种,但这个门与泉古菌门演化关系密切 。其它最近探别出来的古菌与已知古菌门类关系较远,如2006年发现的环境样品 ARMAN ( 英语 : Archaeal Richmond Mine Acidophilic Nanoorganisms ) 。 ARMAN ( 英语 : Archaeal Richmond Mine Acidophilic Nanoorganisms ) 是已知的生命体中最小的生命体。
一个超门(superphylum)分类-TACK,已经被提出,包括 奇古菌门 ( 英语 : Thaumarchaeota ) , 曙古菌门 ( 英语 : Aigarchaeota ) ,泉古菌门和初古菌门 。这个超门可能与真核生物的起源有关。最近,分类Asgard超门名称被命名,并被提议与真核生物的起源和一个TACK姊妹组织更紧密相关 。
细菌、古菌和真核生物的比较
在细胞结构和代谢上,古菌在很多方面接近其它原核生物。然而在基因转录和转译这两个分子生物学的中心过程上,它们并不明显表现出细菌的特征,反而非常接近真核生物。比如,古菌的转译使用真核生物的启动和延伸因子,且转译过程需要真核生物中的TATA框结合蛋白和TFIIB。许多这些特征也被讨论如下。
古菌还具有一些其它特征。与大多数细菌不同,它们只有一层细胞膜而缺少肽聚糖细胞壁。而且,绝大多数细菌和真核生物的细胞膜中的脂类主要由甘油酯组成,而古菌的膜脂由甘油醚构成。这些区别也许是对超高温环境的适应。古菌鞭毛的成分和形成过程也与细菌不同。
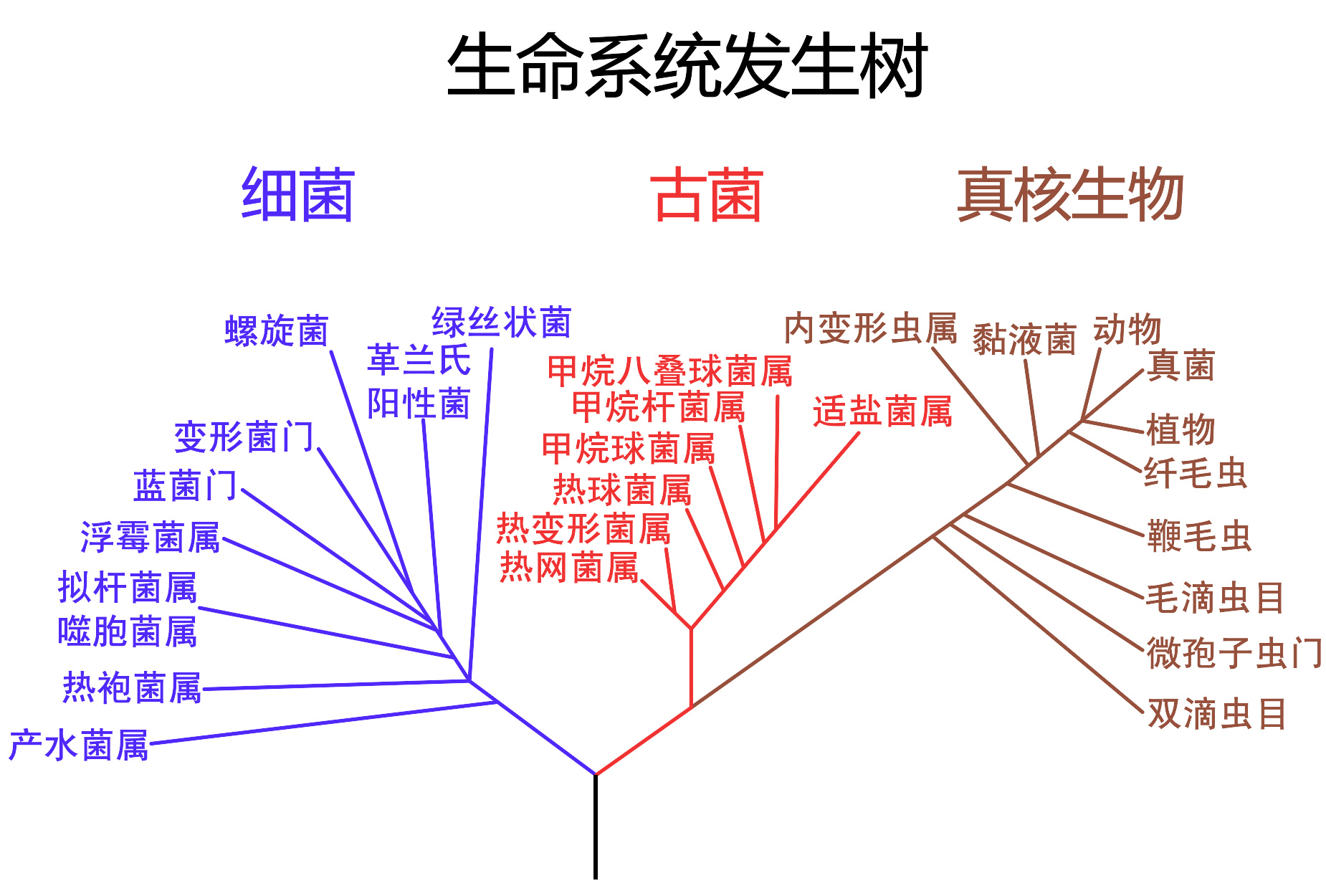
基於rRNA序列的系統發生樹,顯示了可明顯區別的三支:細菌、古菌和真核生物
古菌和真核生物的关系仍然是个重要问题。除掉上面所提到的相似性,很多其他遗传树也将二者并在一起。在一些树中真核生物离广古菌比离泉古菌更近,但生物膜化学的结论相反。然而,在一些细菌,(如栖热袍菌)中发现了和古菌类似的基因,使这些关系变得复杂起来。一些人认为真核生物起源于一个古菌和细菌的融合,二者分别成为细胞核和细胞质。这解释了很多基因上的相似性,但在解释细胞结构上存在困难。
形态
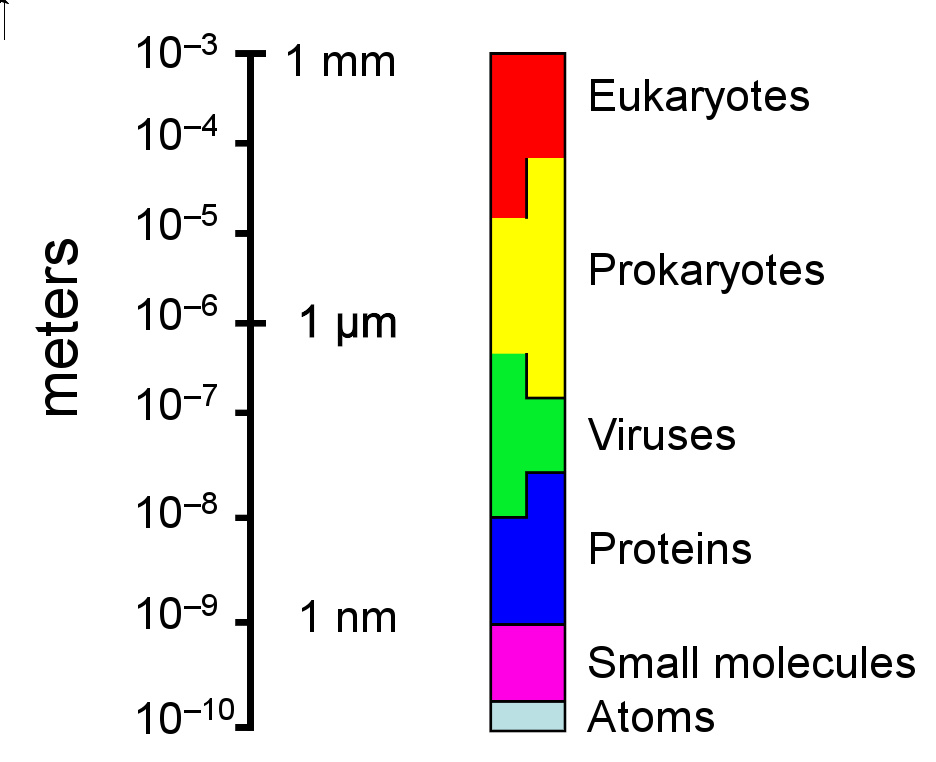
原核细胞的大小,相对于其他细胞和生物分子(对数刻度)
单个古菌细胞直径在0.1到15微米之间,有一些种类形成细胞团簇或者纤维,长度可达200微米。它们可有各种形状,一般是球形、杆形、螺旋形、叶状或方形。这些不寻常的形状可能是由它们的细胞壁和原核生物细胞骨架两者维持的。其他生物的细胞骨架成分蛋白在古菌内存在, 长丝在其细胞内形成, 但与其他生物体不同的是,这些细胞结构被理解的很少。
细胞结构、组成和运行
古菌和细菌的细胞结构大体类似。古菌和细菌一样没有内膜系统和细胞器 。古菌也和细菌一样在细胞膜之外还有一层细胞壁,它们同样也具有一个或多个鞭毛结构 。
膜
古菌膜由从那些在其它生命形式的强烈不同的分子组成,显示出古菌与细菌和真核生物关系都比较遥远。
细胞壁和鞭毛
大多数古菌(但不是Thermoplasma属和Ferroplasma属)具有一个细胞壁。 在大多数古细菌的壁由表面层蛋白质组成,形成一个S层。一个S层是蛋白质分子的刚性阵列覆盖了细胞的外侧(如同锁子甲)。 该层提供化学和物理保护,并能防止大分子接触细胞膜。 与细菌不同,古菌的细胞壁缺少肽聚糖。
古菌鞭毛操作类似细菌鞭毛,它们的长梗是由在座部的旋转马达驱动。然而,古菌鞭毛在组成和发育上有着明显的不同。 这两种类型的鞭毛是从不同的祖先进化而来的。细菌鞭毛与III型分泌系统共享一个共同的祖先, 但是,古菌鞭毛似乎已经从细菌IV型菌的性菌毛进化而来。 对比,细菌鞭毛是空心的,并且是由其子单元从中心孔向上移动到鞭毛的末端装配,但是,古菌鞭毛是由在基座中添加子单元合成。
遗传学
古菌通常拥有单个环状的染色体 ,其大小差异悬殊。目前已知最大的是 Methanosarcina acetivorans ( 英语 : Methanosarcina acetivorans ) 的基因组,达5,751,492个碱基对 ,而除 里奇蒙德矿井嗜酸纳米古生物 ( 英语 : Archaeal Richmond Mine Acidophilic Nanoorganisms ) (ARMAN)外最小的 骑行纳古菌 ( 英语 : Nanoarchaeum equitans ) 只有490,885个碱基对,不到前者的十分之一,据估只能编码537种蛋白质 。一些小型的独立DNA片段质粒也存在于古菌中,可能通过细胞之间的直接接触传递,其过程可能与细菌之间的接合类似 。

DNA病毒STSV1感染的 硫化叶菌 ( 英语 : Sulfolobus ) ,图片下方尺标为1微米
古菌可被双链DNA病毒感染,这些双链DNA病毒形态各异,包括瓶状、杆状、雨滴状等 。这些病毒在一些嗜热菌中研究得最为详致,特别是 硫化叶菌目 ( 英语 : Sulfolobales ) 和 热变形菌目 ( 英语 : Thermoproteales ) 的古菌中 。最近发现了两类可感染古菌的单链DNA病毒:一种是典型的 嗜盐菌多形病毒 ( 英语 : Pleolipoviridae ) ,这病毒感染嗜盐菌 另外一种嗜热的 圈形病毒 ( 英语 : Spiraviridae ) 感染嗜超热菌(这些古菌生长在90到95°C的温度下) 其中后者有已知最大的单链DNA基因组。古菌可以利用针对病毒基因的RNA干扰和DNA重复影响病毒在寄主内的繁殖。
古菌在在基因上与细菌真核生物有许多不同,大约有15%古菌基因是古菌中独有的。这些古菌独有蛋白质的功能大部分还未知 ,其余已知功能的独有蛋白质大部分属于广古菌门中的产甲烷菌。古菌、细菌、真核生物共有的蛋白质都涉及细胞的核心功能,如转录、翻译和核苷酸代谢 。古菌基因组别的特点还有基因的组织常与功能相关(如催化相同代谢途径的酶会组成一个独有的操纵子), 以及不同的tRNA基因之间差异很大,其氨酰-tRNA合成酶之间也是如此 。
较之细菌的转录与翻译,古菌中的这些过程更接近于真核生物,特别是古菌中的RNA聚合酶与核糖体,可以说非常接近于真核生物中的对应物 。尽管古菌中只有一种RNA聚合酶,但其结构与功能与真核生物中最主要的RNA聚合酶Ⅱ非常相似,且其结合到基因启动子上所需的蛋白质组件(通用转录因子)也非常相似 。但是,其它的古菌转录因子却与细菌中的相似 ,转录后修饰也比真核生物中的简单(大部分古菌基因缺少内含子,其内含子主要集中在tRNA和rRNA基因中 ,少数出现在编码蛋白质的基因中 )。
目前已有一百五十余个古菌基因组已经完成了测序,另外许多测序工作正在进行中。
基因的转移与交换
沃氏富盐菌 一种极端嗜盐古菌,可在两个细胞的细胞质之间形成桥梁,可能用于DNA的任意方向的转移 。
当嗜热的古菌 硫化叶菌 ( 英语 : Sulfolobus solfataricus ) 和 嗜酸热硫化叶菌 ( 英语 : Sulfolobus acidocaldarius ) 在DNA突变源,例如紫外线、博来霉素和丝裂霉素的暴露下,不同种类的古菌会出现不同的变化。硫化叶菌的基因变化不会因为其他物理突变源,如酸碱值和温度, 这说明了古菌的进化是来自于DNA的损伤, 这也说明了紫外线导致的细胞进化促使染色体在硫化叶菌之间交换。重组的几率级别比没有经过紫外线照射的古菌多出三级。 这使科学家猜测可能硫化叶菌为了维修它们自己损坏的DNA而与有类似基因的硫化叶菌交换质粒。这反应可能是原始的性交流,这与被经常研究的细菌固定的DNA交换以维修受损DNA类似。
繁殖
更多资料:无性繁殖
古菌利用二分裂、分裂和出芽进行无性繁殖;古菌不会进行减数分裂,所以拥有同样的基因的一个种的古菌可能拥有不同的形态 。古菌的细胞分裂被它们的细胞周期所控制;在细胞的染色体复制并分离后,细胞开始一分为二 。虽说目前古菌中只有硫化叶菌的复制周期被阐明,但是这样的细胞周期大体上类似于在细菌和真核生物的周期。和真核生物一样,古菌的染色体也可以在多个位点(复制起点)开始用DNA聚合酶复制 。然而古菌用于控制细胞分裂的蛋白(例如FtsZ,制造逐渐缩小的“Z环”以帮助细胞质分裂)和分离两个子细胞的隔膜部分,这些与细菌的二分裂相似 。
但与细菌、真核生物不同,目前未发现有古菌进行孢子生殖 。一些 嗜卤盐菌 ( 英语 : Haloarchaea ) 的种类可以进行 表型转换 ( 英语 : Phenotypic switching ) 并生长成为不同的形态。这些形态包括拥有可以防止 渗透压休克 ( 英语 : Osmotic shock ) 出现的厚细胞壁,这使嗜卤盐菌可以在盐度低的水中存活。这些古菌特征不是生殖结构,但是它们可以帮助古菌在新的环境下生存 。
生态学
生活环境
古菌可以在各种各样的栖息地现并且是全球生态重要的一部分 ,古菌占有地球上20%的生物量。 很多古菌是生存在 极端环境 ( 英语 : extreme environment ) 中的,包括最早发现的古菌也是嗜极生物。 一些生存在极高的温度(经常100 °C以上)下,比如间歇泉、石油井或者海底深海热泉中。还有的生存在很冷的环境或者高盐、强酸或强碱性的水中。然而也有些古菌是嗜中性的,能够在沼泽、废水、海洋和土壤中被发现。 很多产甲烷的古菌生存在动物的消化道中,如反刍动物、白蚁或者人类。古菌通常对其它生物无害,且未知有致病古菌。
海洋中浮游生物(浅绿色)的图,古菌是海洋浮游生物中重要的一部分
极端环境下的古菌主要可以分为四种生理群: 嗜盐生物 ( 英语 : halophile ) 、嗜热生物、 嗜碱生物 ( 英语 : alkaliphile ) 及 嗜酸生物 ( 英语 : Acidophile (organisms) ) 。这些不是具体的分类,这些分类也不是互斥的,因此有些古菌有时属于其中的几类中,不过这仍然可以做为分类的起点。
嗜盐类的古菌,生活在高含盐量的环境中,例如盐湖,盐度比嗜盐细菌可以生活的20-25%要高 。嗜热古菌适合生长在超过45 °C(113 °F)的温度,例如热泉中,超嗜热古菌在温度超过80 °C(176 °F)时生长的最好 。古菌Methanopyrus kandleri的116菌种甚至122 °C(252 °F)繁殖,是生物中最高的记录 。
有些古菌可以生长在极酸性或极碱性的环境中 ,例如嗜酸古菌中的Picrophilus torridus可以生长在pH值为0的环境下,相当于1.2M硫酸 。
古菌可以抵抗极端环境的能力也让科学家推测外星生命具有的可能的特质 ,有些极端的环境和火星的环境有些相近。 ,因此推测这些古菌有可能在陨石上,在各行星之间移动 。
近来许多研究发现古菌不只可在高温及中温的环境下生存,也可以在极低温的环境下生存,例如在极地的海洋中就有许多的古菌 ,不过数量更多的是在海洋非极端环境中的古菌,是浮游生物中的微微型浮游生物 。虽然这种古菌数量很多(占纯生物生物量的40%),但其中几乎都无法在实验室隔离的进行 纯培养 ( 英语 : pure culture ) 。因此古菌在海洋生态中的角色是基础的,古菌对整体生物地球化学循环的影响大部分都还不清楚。 有些海洋的泉古菌可以产生硝化作用,因此推测对海洋的氮循环应该有影响 ,不过这些泉古菌也会使用其他的能量来源 。在海床上的沉积物中可以找到大量的古菌,这也是在海底深度超过一米的区域中,在数量上占大多数的生物体 。
在地球的化学循环中的作用
更多资料:生物地质化学循环
古菌参与了地球上碳、氮、硫的循环。虽说这些细胞活动对生态系统来说是十分重要的,但是古菌也可能制造与人为变化类似的环境影响,甚至可能造成环境污染。
古菌也可以进行大部分氮循环中的化学反应。有一些古菌可以从一个生态系统中移除有机氮,这些化学反应包括一些硝酸盐代谢和反硝化反应;其它古菌可以把氮气加入生态形成有机氮,这些反应包括氮吸收和固氮作用。 最近,科学家发现古菌也参与氨的氧化。这些反应在海洋里是特别重要的。 一些古菌也参与土壤里面的氨氧化。这些古菌会制造硝酸盐,而其它微生物则进一步氧化硝酸盐。最后,植物和其它生物会吸收并利用氧化的硝酸盐。
在硫循环中,利用硫化物氧化代谢的古菌可以把硫从岩石中释放出来,并提供给其它生物。但是这些古菌,例如硫化叶菌,会利用硫代谢并最终产出硫酸,所以这种古菌在废弃的矿场中的生长会导致 矿渣酸性废水 ( 英语 : acid mine drainage ) 的出现以及其它环境污染。
在碳循环中,制造甲烷的古菌可以移除多余的氢并且在有机化合物的缺氧分解中扮演着分解者的角色。这些古菌可以在沉积物和沼泽等缺氧环境中生活,它们也可以被用于污水处理
产甲烷菌是大气甲烷的主要来源,它们每年释放的甲烷占了世界上大部分的甲烷排放。 产甲烷菌排放的甲烷加速了全球的温室气体排放和全球变暖。
全球甲烷数值于前工业化时期的722PPB(10亿吨大气内有722吨甲烷)增加到2011年的1800PPB,这期间甲烷数值上升到了之前的2.5倍,2011年是在80万年内全球甲烷数值最高的一年。 甲烷的全球变暖潜能值是29,这说明了甲烷可以在100年内吸收比二氧化碳多29倍的热量。
与其它生物的关系
更多资料:种间关系

制造甲烷的古菌与白蚁拥有互利共生关系
古菌与其它生物之间的关系主要都是互利共生或者偏利共生。至今还没有发现古菌是任何其它生物的病原体或寄生虫。 虽说至今没有发现并确定古菌病原体,但是研究证明一些产甲烷菌的种类可能与牙周疾病有关。 另外,一种 骑行纳古菌 ( 英语 : Nanoarchaeum equitans ) 可能是泉古菌 Ignicoccus hospitalis 寄生虫,古菌必须在寄主里面生活并繁殖, 但是对寄主没有任何好处。 相反的, 里奇蒙德矿井嗜酸纳米古生物 ( 英语 : Archaeal Richmond Mine Acidophilic Nanoorganisms ) (ARMAN)只是与酸性矿井内生物薄膜的其它古菌偶尔接触。 这种关系的性质至今仍然是未知的。但是科学家知道 Nanarchaeaum 和 Ignicoccus 之间的关系与微小的ARMAN古菌不同,ARMAN古菌一般与 嗜酸热菌 ( 英语 : Thermoplasmatales ) 细胞不会相互影响。
互利共生
所有反刍动物和白蚁的消化道中有其中一种在 产甲烷菌 ( 英语 : methanogen ) 和原生动物之间的互利共生,它们帮助寄主消化纤维素。 在这缺氧的环境中,原生动物利用纤维素获得能量并同时释放产物氢气。大量的氢气会影响原生动物获得能量的效率。当产甲烷菌把氢气经过化学反应变成甲烷后,原生动物会因为能获得更多能量而获益。
一些古菌生活在厌氧原生动物里面并利用寄主 造氢体 ( 英语 : hydrogenosome ) 制造的氢气,这些原生动物包括纤毛虫 Plagiopyla frontata 。 古菌也与更大的生物有共生关系。例如海里的古菌 Cenarchaeum symbiosum 生活在海绵 Axinella mexicana 的里并与海绵拥有共生关系。
片利共生
古菌也可以是片利共生中的获益者,同时寄主既没有获益也没有受害。例如 史密斯甲烷菌 ( 英语 : Methanobrevibacter smithii ) 就是在的人体正常肠道菌群中最常见的古菌,这种古菌占了肠道中原核生物的十分之一 。在白蚁和人体内,这些产甲烷菌甚至可能是有益的,这些古菌会与其它菌群互相影响并协助消化 。古菌的菌落也与不少其它生物表面或附近生活,例如在珊瑚的表面 和环绕植物根冠的根际。
在技术与工业中的作用
更多资料:生物技术
一些耐热或耐酸碱的嗜极古菌可以作为一些在极端环境下活跃的酶的来源。 这些来自古菌的酶拥有很多用途。其中耐高温的DNA聚合酶,例如从 焦酚火球菌 ( 英语 : Pyrococcus furiosus ) 中提取的 火球菌DNA聚合酶 ( 英语 : Pfu DNA polymerase ) 可以在需要进行聚合酶链式反应时简单并快速地复制DNA,这彻底改变了对分子生物学地研究。在工业中,来自其它 焦酚菌 ( 英语 : Pyrococcus ) 的淀粉酶、 半乳糖苷酶 ( 英语 : galactosidases ) 和 普鲁兰酶 ( 英语 : pullulanase ) 可以在高达100 °C(212 °F)的环境下仍然活跃,这使食品加工可以在高温下进行,这些酶可以被用来加工低乳糖牛奶和乳清。 嗜热古菌生产的酶可以在有机溶剂内仍然保持稳定,这些酶可以在保持环保的前提下用来合成有机化合物。 这种稳定性使它们更容易在结构生物学中研究。这就是为什么与细菌和真核生物对应的嗜极古菌生产的酶经常被用于分子结构的研究。
相较于酵素领域的应用,古菌本身在生物科技上的应用就比较少了。产甲烷的古菌是进行厌氧消化,产生生物气体的微生物群体的一部分,是污水处理中重要的一环 。在矿石处理中,嗜酸古菌可以用来从矿物中萃取包括金、钴及铜等金属 。
古菌也是一些潜在有效抗细菌药的来源,已经分析了其中几种的古菌素,一般认为有几百种,特别是由 嗜盐细菌 ( 英语 : Haloarchaea ) 及 硫化叶菌 ( 英语 : Sulfolobus ) 所产生的。这些古菌素的结构和从细菌产生的抗细菌药不同,所以可能有新的作用方式。此外它们可以创建新的 选择标记 ( 英语 : selectable marker ) ,可以用在古菌分子生物学中 。
新分类
“Miscellaneous Crenarchaeota Group” (MCG)古菌是迄今为止发现分布最为广泛的一类未培养古菌,不仅分布广泛,而且数量庞大,被认为是海底深部生物圈中最丰富,并且最活跃的类群之一,很可能在全球物质和能量循环过程中发挥了重要的作用。然而到目前为止,尚未获得此类古菌的纯培养,其生理和功能特征以及在环境中发挥的作用都不得而知,并且其分类地位也不明确,暂时归类于泉古菌门。研究者们发现MCG古菌在深部生物圈沉积物中广泛并大量存在,代表了一类自然界较古老的古菌,在系统发育上处于一个深的分支,显著不同于目前分类已确定的所有古菌门类,包括新近确立的奇古菌门和曙古菌门。因此,提议将MCG古菌归类于一个全新的门类,命名为深古菌门(Bathyarchaeota)。是古菌和生命起源和演化研究的重要进展之一,也将为本领域的科学研究起到积极推动作用。
参见
古菌分类表
已测序古菌基因组列表
古菌属列表
参考文献
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。



- 有价值
- 一般般
- 没价值
















推荐阅读


关于我们

APP下载


