






人类演化
思想史
人属的英文 homo 这个词,这个用来称呼人类所属的生物属名,是拉丁文称呼“人类”的字词。它最初是由卡尔·林奈选来用在他的分类系统中。英文的“人类”一字human源自于拉丁文的 humanus ,这是 homo 的形容词。拉丁文的 homo 源自于印欧语系的语根 dhghem ,意指“大地”。
卡尔·林奈及当时其他科学家也认为,大猿是人类最亲密的近亲,这是由于形态学和解剖上的相似处。人类与早期类人猿在谱系上具有连结的这种可能性,直到1859年查尔斯·达尔文出版《物种起源》之后,才可能变得更清楚。这本书主张支持新物种由早先物种演化而来的概念。达尔文的书没有触及人类演化的问题,只是说:“光芒将会投射在人类起源和他的历史”。

在美国俄克拉荷马州人骨学博物馆(The Museum of Osteology)的人科动物化石演化展示
最早关于人类演化本质的争辩,出现在汤玛斯·亨利·赫胥黎和理查·欧文之间。赫胥黎认为人类从类人猿演化而来,他列举了人类和类人猿之间的许多相似性与相异性,特别是在他1863年的书《有关人类在自然界定位的证据》( Evidence as to Man"s Place in Nature )。然而,许多达尔文的早期支持者(如阿弗雷德·罗素·华莱士和查理斯·莱尔)并不同意人类心智能力和道德情感的起源,可借由自然选择得到解释。达尔文于1871年发表《人类的由来》一书时,将演化理论与性选择运用于人类本身。
有一个严重的问题,就是缺乏介于人类与类人猿中间类型的化石。直到1920年代,这类化石才在非洲出土。1925年,雷蒙·达特(Raymond Dart)描述了南方古猿非洲种( Australopithecus africanus )的命名用标本(type specimen)是“陶格孩童”(Taung Child),这是在一座山洞里发现的一具南方猿人婴孩。这个孩童的遗骸包括一个保存完好的细小头骨及其脑子的颅内腔模型(endocranial cast)。虽然这个脑子的脑容量较小(410立方公分),但是它的形状是圆转的,有别于黑猩猩和大猩猩的脑子,而更像是现代人类的脑子。此外,这个标本也显示较短的犬齿,而且枕骨大孔的位置成为双足直立行走的证据。所有这些特征都使得达特相信,陶格孩童是一个双足直立行走的人类祖先,也是介于类人猿与人类之间的一个过渡形式。
类人猿分类学(人类及其近亲的分类)自从1950年代以来,已有很大的变化。 例如,纤细种南方猿人曾被认为是现代人所属的人属( Homo )祖先。 南方古猿与智人都是动物分类学上的人族(tribe Hominini)的一部分。
1970年代所搜集的资料显示,南方古猿是个多样化的群体,南方古猿非洲种未必是现代人类的直接祖先。 原先将南方猿人分类为“纤细种”与“粗壮种”,现在的重新分类将粗壮种放进单独的属:傍人( Paranthropus )。 分类学家将人类、南方古猿及相关物种,以及其他类人猿放在同一个科,称为人科(Hominidae)。
在1980年代与1990年代DNA测序的进展,特别是线粒体DNA以及稍后的Y染色体DNA的研究,增进了对人类起源的认识。 来自世界各地广大原住民群体的线粒体DNA与Y染色体DNA样本排序结果,显示了关于男性与女性基因遗传的远祖讯息。 基因树状图差异的校准,支持了现代人类晚近非洲起源说。 分析呈现了在非洲各地具有最大的DNA模式多样性,这显示非洲是线粒体夏娃与Y染色体亚当的祖居地。
在人属出现之前
大猿的演化
更猴
灵长类的演化史可追溯到距今6500万年,它们是在当时造成物种(特别是恐龙)灭绝的大灾难下幸存的胎盘哺乳动物之中,最古老的一群。已知最古老的类灵长类哺乳动物物种,更猴(Plesiadapis)来自北美,但它们遍布于欧亚大陆和非洲,在古新世和始新世期间,这两处地方都处于热带。

假熊猴
随着当代气候的开始,特点是在大约距今3000万年前的渐新世第一次出现了南极冰帽。在这个时间出现的灵长类动物是假熊猴(Notharctus)。1980年代在德国发现的化石证据,并正在挑战关于人类祖先源自非洲大陆的理论。
大卫·毕甘(David Begun) 表示,这些灵长类动物在欧亚大陆蓬勃发展,而这个谱系导向非洲猿类和人类,包括森林古猿(Dryopithecus),从欧洲或西亚向南迁移至非洲。这支后来存活下来的热带群体,出土最完整的一群见于开罗西南部的法扬(Fayum)堆积层,始新世晚期和渐新世最早期的化石层位之中,这引发后续所有的现生灵长类,马达加斯加的狐猴,东南亚的懒猴,非洲的婴猴属(galago)或“丛林婴儿”(bush babies),以及人形亚目(Simian anthropoids);广鼻猴次目(platyrrhine)或新世界猴,和狭鼻猴次目(catarrhine)或旧大陆猴、类人猿和人类。
已知最早的狭鼻猴次目,来自渐新世晚期的卡莫亚猿( Kamoyapithecus ),在肯亚北部大裂谷的Eragaleit出土,定年为距今2400万年前。其祖先一般认定是来自法扬的埃及猿、原上猿与傍猿( Parapithecus ),在大约距今3500万年前。在2010年, Saadanius 被描述为狭鼻猴次目冠状群体(Crown group)最后共同祖先的一个近亲,目前暂时定年到距今2900万到2800万年间,有助于填补长达1100万年的化石记录断层。

经过重建的无尾的原康修尔猿骨架
在中新世早期,大约2200万年前,来自东非的多种适应树居生活的原始狭鼻猴亚目,显示一段历史悠久的早期多样化发展。在2000万年前的化石,包括归类于最早的旧世界猴维多利亚猿( Victoriapithecus )的碎片。在这个属当中,被认为处在猿类谱系之内,可推至距今1300万年前的物种包括原康修尔猿、 Rangwapithecus 、 Dendropithecus 、 Limnopithecus 、 Nacholapithecus 、 Equatorius 、 Nyanzapithecus 、非洲古猿、日猿( Heliopithecus )和肯尼亚古猿,都来自东非。其他中新世的普遍非cercopithecids来自遥远的遗址,沉积在纳米比亚洞穴的 Otavipithecus ,出自法国、西班牙和奥地利的加泰罗尼亚皮尔劳尔猿和森林古猿,证实在中新世早中期,在气候相对温暖的非洲和地中海盆地,有着广大的生物型态多样性。年代最近的中新世类人猿,山猿,出土自意大利距今900万年的煤层。
分子证据指出,长臂猿(长臂猿科)的谱系,大约在距今1800万至1200万年前,从大猿的谱系分割出来;红毛猩猩(猩猩亚科)的谱系,大约在距今1200万年前,从大猿的谱系分割出来。目前并未出土足以清楚记载长臂猿祖先的化石,它可能源自于一个迄今未知的东南亚类人猿群体,但化石原红毛猩猩可由来自印度的拉玛古猿( Ramapithecus )和来自土耳其的 Griphopithecus 做为其祖先的代表,追溯至距今1000万年前。
人类谱系从其他大猿分裂两族和之后

一个雌性的阿法南方古猿(南方猿人阿法种)的重建模型。绰号“露西”。
最接近大猩猩、黑猩猩和人类最后的共同祖先,可由肯亚出土的仲山纳卡里猿和希腊出土的欧兰猿为代表。分子证据显示,在距今800万至400万年前,首先是大猩猩,然后是黑猩猩,从这条通往人类的演化路线分离出来;人类的DNA大约有98.4%与黑猩猩相同,这是比较单核苷酸多态性(single nucleotide polymorphisms)所得到的结果。大猩猩和黑猩猩的化石记录相当有限。两者的保存状况都不理想(热带雨林土壤往往是酸性,会分解骨骼)。
其他人族有可能适应赤道带以外的干燥环境,还有羚羊、土狼、狗、猪、大象以及马。约在距今800万年前,赤道带缩小。这些化石人族–从与黑猩猩分道扬镳之后的人类谱系 - 比较广为人知。最早的是乍得沙赫人(距今700万年,乍得沙赫人可能是人与黑猩猩的最近共同祖先)和图根原人(距今600万年),其次是:
地猿(距今550万到440万年),包括地猿始祖种( Ardipithecus kadabba )和地猿根源种( Ardipithecus ramidus )。
南方古猿属( Australopithecus ,距今400万到180万年),包括湖畔南方古猿、阿法南方古猿、南方古猿非洲种、羚羊河南方古猿、惊奇南方古猿,以及源泉南方古猿。
肯亚猿人属("Kenyanthropus ,距今300万到270万年)。
傍人属(距今300万到120万年),包括埃塞俄比亚傍人、鲍氏傍人、粗壮种傍人( Paranthropus robustus )。
人属(距今200万年至今),包括巧人、人属鲁道夫种、匠人、格鲁及亚人、人属先驱种、西布兰诺人、直立人、海德堡人、罗德西亚人、尼安德塔人、智人伊达图亚种、古智人、弗洛瑞斯人。
人属
智人是人属唯一的现生物种,另有某些已告灭绝的人属物种可能是智人的祖先,有许多可能是我们的“表兄弟”,已经透过物种生成过程而分离出去(speciated),远离我们祖先的路线。 关于哪些群体应算作单独的物种,哪些是亚种,目前尚未达成共识。在某些情况下,这是由于缺乏化石证据;在其他情况下,这是由于使用细微的差异,来分类人属的各种不同物种的结果。“撒哈拉泵浦理论”(Sahara pump theory) (描述一个偶尔勉强称得上“湿润”的撒哈拉沙漠)提供对早期人属变异的解释。
依据考古学和古生物的证据,有可能在某种程度上推断出,各种古代人属的饮食习惯,并研究在人属之中,饮食对于体质与行为演化所扮演的角色。
巧人

巧人的重建模型。
巧人( H. habilis )存活在大约距今240万到140万年前。巧人是人属的第一个物种,在上新世晚期或更新世早期的非洲南部与东非,大约距今250万到200万年前,从南方猿人分支出来。巧人具有比南方猿人更小的大臼齿与更大的脑子,并且运用石头(可能也用动物骨骼)制作工具。这是目前已知最早的人属动物之一,其发现者路易·李基(Louis Leakey),因着它与石器的关连性,昵称为“巧人”。某些科学家提议将这个物种的属别,从人属移到南方猿人属,因为依据对于它的骨架所做的型态学分析,它比较适应于树居(arboreal,栖息在树上),而不是如同智人一般双足直立行走(bipedalism)。
人属鲁道夫种与人属乔治亚种
这些物种名称是用在大约从距今190到160万年间的化石,然而它们与巧人的关系尚未厘清。
卢多尔夫人( Homo rudolfensis )指称来自肯尼亚的一个不完整的头骨。科学家建议这是另一个巧人,但这尚未获得确认。
格鲁及亚人( Homo georgicus )来自格鲁吉亚共和国,可能是介于巧人与直立人间的一个中间型态物种, 或是直立人的一个亚种。
匠人与直立人
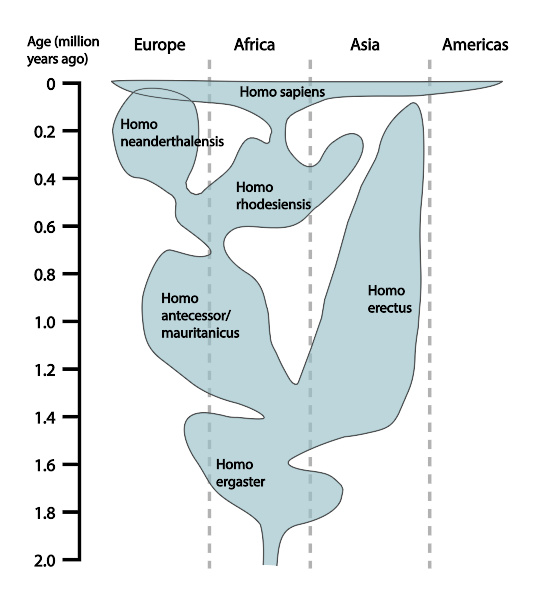
关于人科群体基于化石证据的时空分布模拟, 其他诠释方式的主要差别在于人科物种的分类学与地理分布。
第一个直立人化石是由荷兰军医杜保斯(Eugene Dubois)在1891年于印度尼西亚的爪哇岛发现的。他起初将这批化石材料命名为 Pithecanthropus erectus ,这是依据这些化石的型态学,他认为这是介于人类与大猿的中间物种。 直立人( Homo erectus )存活的期间大约从距今180万年到7万年前(这或许指出,它们可能受到距今约7万年前的多峇巨灾(Toba catastrophe)影响而灭绝;然而,直立人梭罗亚种( Homo erectus soloensis )与弗洛瑞斯人( Homo floresiensis )逃过这场巨灾而存活下来。在前半段,从距今180万到125万年前的直立人,被认为是一个独立的物种:匠人( Homo ergaster ),或是直立人的亚种:直立人匠人亚种( Homo erectus ergaster )。
在更新世早期,距今150万到100万年间的非洲、亚洲与欧洲,某些巧人群体发展出较大的脑子,并制作更进步的石器;连同其他差异,足以让人类学家将它们分类为一个新物种:直立人。此外,直立人确实是第一个直立行走的人类祖先。 这可能借由膝盖的演化,以及枕骨大孔(foramen magnum,人类头骨正下方的大孔,连接脑子与脊髓)的位置前移而成为可能。直立人用火煮食肉类。
直立人的有名标本是北京人(因其出土化石数量庞大且层位较多);其他标本在亚洲(特别在印度尼西亚)、非洲与欧洲出土。现在有许多古人类学家用“匠人”来指称这个群体的非亚洲型态,“直立人”这个名称只保留用在亚洲所发现的化石,以呼应它们在骨架与齿式方面有别于“匠人”之处。
西布兰诺人与人属先驱种
这些物种可能是介于直立人与人属海德堡种之间的中介物种。
人属先驱种( Homo antecessor )已知是来自西班牙与英国的化石,定年为距今120万年到50万年前。
西布兰诺人( Homo cepranensis )指称一个在意大利出土的头盖骨,据估计约在距今80万年前。
人属海德堡种
人属海德堡种( H. heidelbergensis )存活在大约80万到30万年前。学者也提议将其分类为智人海德堡亚种( Homo sapiens heidelbergensis )或智人古匈牙利亚种( Homo sapiens paleohungaricus )。
罗德西亚人与贾威斯头骨
罗德西亚人( H. rhodesiensis ),估计其年代约在距今30万年到125,000年前。目前大多数的专家相信罗德西亚人是属于人属海德堡种,即使其他学者提议其为古智人与智人罗德西亚亚种( Homo sapiens rhodesiensis )。
在2006年2月,贾威斯头骨(Gawis cranium)的化石出土,这可能是介于直立人与智人之间的中间物种,或是许多已告灭绝的演化路线之一。这个来自埃塞俄比亚贾威斯的头骨的年代约在距今50万年到25万年间。目前只有出版摘要资料,尚未由这个团队发表经过同僚审查的研究报告。贾威斯头骨的脸部特征显示,它是一个中间物种或是“波多人”(Bodo man)女性的标本。
尼安德塔人
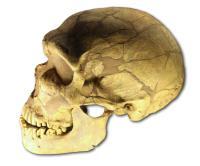
Le Ferrassie的尼安德塔人头骨 (模型)
尼安德塔人( H. neanderthalensis )存活的时间从距今40万年前。 也有学者提议将其分类为智人尼安德塔亚种( Homo sapiens neanderthalensis ):学者持续争辩“尼安德塔人”是否为一个单独物种:人属尼安德塔种,或者是智人的一个亚种。 虽然这个争论尚未平息,来自线粒体DNA排序的证据指出,在尼安德塔人与智人之间并未发生明显的基因流布。因此,这两者是分别的物种,其共同祖先约在距今66万年前。 在1997年,马克史东庆(Mark Stoneking)说道:“[依据从尼安德塔人骨骼所萃取的线粒体DNA]得到的结果指出,尼安德塔人并未贡献线粒体DNA给现代人类…尼安德塔人并不是我们的祖先”。对于第二组尼安德塔人来源的调查,支持了这些发现。 然而,在2010对于尼安德塔人基因体的排序指出,尼安德塔人确实在大约公元前75,000年与智人混血(在智人迁移离开非洲之后,但在他们分别迁往欧洲、中东与亚洲之前)。 当代人类的DNA约有1%到4%源自尼安德塔人的DNA。 这个1-4%的DNA差异只出现在非洲以外的人类。 然而,“多区域演化”假说的支持者指出,最近研究显示,非洲以外的核心DNA遗传的时间可回溯到距今100万年, 即使这些研究的可靠性受到质疑。 来自智人的竞争,可能导致尼安德塔人的灭绝。 他们有可能在欧洲并存,直到距今1万年前。
智人
智人( Homo sapiens ,种名 sapiens ( 英语 : wikt:sapiens ) 的拉丁文原意为“聪明”或“有知识”)存活于距今25万年到现在。在大约距今40万年至25万年间,当时是更新世第二间冰期,人类的脑容量变大,且发展出更细致的石器制作技术,这些都是由直立人转变为智人的证据。直接的证据显示,有一股直立人从非洲向外迁移,接着在非洲的物种生成过程中,智人从直立人演化出来。智人后续在非洲内部与向非洲以外地区的迁移,最终取代了较早散居各地的直立人。这个迁移与起源理论往往称为单地起源说或非洲起源说。目前的证据并未排除某些多区域演化(multiregional evolution)或迁居的智人与各地既有的人属群体混血的可能。这是目前古人类学的一个热烈争辩的领域。
当前的研究确认了,人类在遗传学上是高度同质的;换言之,人类个体的DNA比起其他物种而言,具有较多的相似性。这可能源自人类演化的时间相对晚近,或由某些毁灭性的自然灾害所引起的种群瓶颈,例如多峇巨灾。 然而,人类各群体已产生独特的遗传特征,主要导因于人类的小群体迁移到许多新环境。这些因适应而产生的特征,仅占智人基因体的极小成分,但包括了各种不同特征,例如肤色与鼻型,此外也有内部构造的特色,例如在高海拔地区更有效呼吸的能力。
来自埃塞俄比亚的长者智人可能是一个已消失的亚种,存活于大约距今16万年前。
弗洛瑞斯人
File:Homo floresiensis - reconstruction.JPG 弗洛瑞斯人女性头部的重建模型
弗洛瑞斯人( H. floresiensis )大约存活在距今10万年到12,000年间,由于体型娇小,被昵称为哈比人,这可能是岛内侏儒化所造成的结果。 弗洛瑞斯人引人好奇的是它们的体型与年代,这是一个具体的晚近的人属物种,但展现了许多当代人类所没有的衍生特征(derived traits)。换言之,弗洛瑞斯人与当代人类具有共同祖先,但从当代人类的谱系分出去,依循一条独特的演化路径。主要的发现是一个年约30岁的女性骨架。在2003年出土,定年约在距今18,000年。这名女子存活时身高约一米,脑容量约380立方公分。(其脑容量如同黑猩猩,不到智人的平均值1400立方公分的三分之一。)
然而,学界持续争辩著,究竟“弗洛瑞斯人”是否确实是个单独存在的物种。 目前有一些科学家相信,“弗洛瑞斯人”是受到病理侏儒症所苦的一种“智人”。 这一假说受到部分的支持,因为在弗洛瑞斯岛,也就是这个骨架被发现的岛屿,有某些当代人类是俾格米人。有人认为,这些较矮的俾格米人再加上病理侏儒症,确实可能创造出一种类似哈比人的人类。其他主要对于“弗洛瑞斯人”这个物种概念所做的攻击,在于在其出土地点的工具仅仅与智人有关。
但是,这个病理侏儒症的假说无法解释的是,为何弗洛瑞斯人其他的解剖特征并不像现代人类(无论患病与否),反而很像我们种属的古代成员。除了头骨特征之外,这些特征也包括手腕、前臂、肩膀、膝盖和脚的骨骼形式。
丹尼索瓦人族
2008年,在西伯利亚阿尔泰山从事研究工作的考古学家,于丹尼索瓦洞穴发现一个小指骨头碎片,来自目前称为丹尼索瓦人(Denisova hominin)群体的一位小女孩。 在这处洞穴的相同层位出土的器物,包括一个手镯,依据放射性碳定年法大约在距今四万年前。由于丹尼索瓦洞穴气候凉爽,因此在化石片段之中的DNA得以保存下来,目前已对其线粒体DNA及核心基因组DNA完成排序。
虽然其线粒体DNA的分歧点时间深度远比预期更深, 但是完整的基因组排序,显示丹尼索瓦人与尼安德塔人属于相同的世系,但两者就在现代人类兴起不久之后,就从同一条路线分裂开来。 目前已知在欧洲,现代人类与尼安德塔人的重叠时间超过一万年,而且这项发现引发一种可能性,尼安德塔人、现代人类、丹尼索瓦人或许曾经并存。帕博(Pääbo)指出,这个分支的存在,使得更新世晚期的图像更加复杂。
人属物种对照表
使用工具

“尖石”,这是一个奥度瓦伊砾石器,人类石器的最基本类型。

火是人类最伟大的发现之一,对于人类演化非常重要。

从英国肯特郡出土的Acheulean手斧,这是直立人使用的燧石器。

维伦多尔夫的维纳斯,旧石器时代艺术的范例。
使用工具已被学者诠释成一种智力的标志,而且学者提出理论,工具使用可能触发了人类演化的某些层面----最值得一提的是人类脑子的持续扩张。古生物学目前依然未能解释脑子经过数百万年的扩张,即使说从能量消耗来看,脑子的需求非常大。当代人类的脑子消耗约20瓦(每天400千卡),占人体能量消耗的五分之一。工具使用的增加,使得人类能猎得能量丰富的肉类,并有能力处理更多能量丰富的植物产品。研究人员认为,早期人类是在演化压力下,提高他们创造和使用工具的能力。
值得一提的是,有许多物种能够制作和使用工具,但人类这个物种控制了制作的范围并使用更复杂的工具。目前已知最古老的工具来自埃塞俄比亚的“奥度瓦伊石器”(Oldowan Stone Tools,发现地点为奥度瓦伊Olduwai)。这些工具的年代距今250万年到260万年,来自已知最早的“人属”物种。目前尚无证据显示,距今250万年前有任何“人属”标本。在一些奥度瓦伊工具附近,出土了一个人属化石,值得注意的是其定年在距今230万年,这隐约显示,也许早期人属物种确实创造和使用这些工具。这确实可能,但没有坚实的证据。伯纳伍德(Bernard Wood)指出,“傍人属”与这个“奥度瓦伊技术丛”("Oldowan Industrial Complex")大致同时存在这个区域。虽然没有直接证据可佐证傍人属是工具制造者,但他们的解剖构造提供了间接证据,显示他们具备这方面的能力。大多数古人类学家都同意,确实最有可能是早期的“人属”物种制作出奥度瓦伊工具。他们认为,当大多数的奥度瓦伊工具与人类化石同时被发现时,出现的始终是人属,但不是傍人属。
1994年,兰德尔·苏斯曼采用人属及傍人属的对向性拇指来做为他的立论基础,推论他们都是工具制造者。他比较了人类和黑猩猩的拇指骨骼和肌肉,发现人类具有三条黑猩猩所欠缺的肌肉。人类也有较厚实的掌骨与更大的头部,这使得人类的手在精准抓握方面,比黑猩猩更成功。苏斯曼提出辩护,人类拇指的现代解剖学构造是基于制作与操作工具的需求而产生的演化回应,这两个物种确实是工具制造者。
石器
经证实,最早的石器约在距今260万年前,当时在东非的巧人使用的工具,是由圆形砾石简单捶打而成的砾石器、砍器。 这标示了旧石器时代的开始,结束在最后一次冰河时代告终之时,大约距今一万年前。旧石器时代细分为旧石器时代早期(约在距今35万至30万年前结束),旧石器时代中期(直到距今五万年到三万年前)和旧石器时代晚期。
从距今70万至30万年的这段时期,称为阿歇尔传统〈Acheulean〉,当时的匠人(或直立人)由燧石和石英石制作手斧,起初相当粗糙(早期阿歇尔),后来经过在石片两侧更多细部修整。在距今35万年之后,更细致的勒伐技术(Levallois)发展出来。它包含一系列连续敲击,制作出刮削器、切割器、针、扁针。 最后,大约在距今五万年之后,尼安德塔人及克罗马侬人制作出更细腻与专门化的燧石工具(刀、石瓣器)。在此期间,他们开始制作骨制工具。
朝向行为现代性的转型
直到大约距今5万到4万年前,石器的使用似乎逐步取得进展。每个阶段(巧人、匠人、尼安德塔人)从比起前一个阶段更高的层次开始,但每个阶段开始后,进一步发展就迟缓下来。当前的古人类学家,正在争辩这些智人物种究竟是否具备跟现代人有关的某些或许多文化和行为特征,诸如语言、复杂的象征思维、科技创造力等等。看起来,他们有很长一段时间,是维持简单的技术和觅食模式的文化保守分子。
大约距今5万年前,当代人类文化开始更快速地演进。这项朝向行为现代性的转变,称为欧亚大陆人的“”, 或称为“旧石器晚期革命”, 这是由于在考古学记录中,突然大量出现当代行为的迹象。某些学者认为这个转型是更为渐进的,花费更长时间,因为有些特征早在距今20万年前的古非洲智人当中。
现代人类开始埋葬死者、制作衣服、发展复杂的狩猎技巧(如使用诱捕坑或逼使动物跳下峭壁),并从事石洞壁画。 随着人类文化的进展,人类的不同人群推出新颖的技术,加入既有的技术当中:鱼钩、钮扣与骨针等等的器物,呈现了不同人类群体之间的变异,某些东西未曾出现在距今5万年前的人类文化当中。一般来说,尼安德塔人的各个群体在技术方面并未有所差异。
人类学家指出的现代人类行为的具体例证,包括专业化的工具、珠宝首饰和图像的使用(如洞穴壁画)、生存空间的组织、仪式(例如,有陪葬品的墓葬)、专门的狩猎技术、探索较不适合人居的地理区域、以物易物的贸易网络等等。学者之间的争辩还在持续进行,围绕在究竟是一场“革命”导致现代人类的产生,或者说这个演变是渐进的。
人类演化的模型
今天,所有的人类都同属单一的群体:智人智人亚种( Homo sapiens sapiens ),并没有物种区隔。然而,根据“源出非洲”模型,这并不是人属的第一个物种:人属的第一个物种“巧人”,在至少距今两百万年演变出来,巧人生活在非洲不同区域的时间相对较短,“直立人”在超过距今180万年前演变出来,到了距今150万年前就已散布到旧世界各地。
人类学家对于当代人类群体演化有着分歧的看法,是否只在东非发生演变、物种形成,然后迁移到非洲以外,并取代欧亚大陆的人类群体(所谓的“源出非洲”模型或“完全替代”模型);或是演变成一个相互连结的群体(由“多区域演化”假设所假定的模型)。
源出非洲
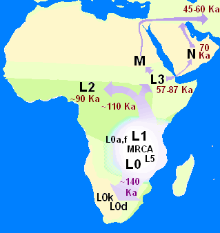
线粒体DNA的分支,这种 DNA只透过母亲传给女儿。
依据由克里斯·斯特林格和彼得·安德鲁斯所发展的源出非洲模型(Out-of-Africa model),现代智人是在距今20万年前的非洲演变而来的。智人大约在距今七万年到五万年间,开始从非洲迁移出来,并最终取代当时存在于欧洲和亚洲的原始人属物种。 源出非洲已获得研究的支持,包括对女性的线粒体DNA(mtDNA)和男性Y染色体的研究。使用133个类型的线粒体DNA构建谱系树状分析后,研究人员得出结论,她们都是来自非洲的同一个女人的后代,这个女人称为线粒体夏娃。源出非洲模型也从非洲人群具有最高的线粒体遗传多样性这项事实得到支持。
关于这个向外迁徙的过程究竟是一次或多次,学者抱持着不同的理论。一套多次迁徙模型,其中包括向南扩散理论, 近年来已经获得来自基因、语言和考古证据的支持。在这个理论中,大约在距今七万年前,现代人类从非洲之角展开一场沿着海岸的迁徙。这群人有助于将人类扩展到东南亚和大洋洲,这解释了为何早期人类在这些区域的遗址年代,远远早于在黎凡特的遗址年代。人类的第二波迁徙浪潮跨越了西奈半岛进入亚洲,产生了在欧亚大陆的大批人群。第二波的人群拥有更成熟的工具技术,而且比起第一波的人群更少依赖沿海的食物来源。第一波人群的扩张证据可能已被每一次冰河时期结束时,上升的海平面所摧毁。 这个多次迁徙模型与来自线粒体DNA的研究结果相冲突,后者显示欧亚大陆的人群以及东南亚与大洋洲的人群都是从同一个线粒体DNA谱系传承下来,这支持了单一的源出非洲迁徙过程,产生了所有在非洲以外的人类群体。
莎拉·提胥可夫率领的广大非洲的遗传多样性研究团队,从113个不同人群的采样当中,发现布希曼人具有最大的遗传多样性,这使得他们成为14个 人类基因丛集 ( 英语 : Human genetic clustering ) 的其中一个。这项研究也定位了非洲西南部现代人类迁移的源头,接近纳米比亚和安哥拉海岸。
根据人类学家和考古学家所采用的多峇巨灾理论,大约在距今七万年前,印尼苏门答腊岛的多峇湖发生一场超级火山喷发,造成了全球性的后果, 致使当时的大多数人死亡,并创造出一个种群瓶颈,影响现今所有人类的基因遗传。
多地起源模型
“多地起源模型”是一种“解释人类演化的模型”,由沃波夫(Milford H. Wolpoff)于1988年提出。 多区域演化的观点主张,人类演化是从距今250万年前的更新世开始,直到现在,一直维持着单一的、连续的人类物种,在世界各地演化成为现代智人。依据多区域假设,化石及基因体资料是说明全球性人类演化的证据,这与最近由晚近非洲起源说所假定的物种形成过程相矛盾。理查李基(Richard Leakey)认为化石证据并不充足,无法解决这个争论。 对于Y染色体及线粒体DNA的单倍群研究已大大支持一个晚近的非洲起源。 来自常染色体DNA的证据也支持晚近非洲起源的可能性。然而,某些研究显示,古智人(Archaic Homo sapiens)的遗传成分也有可能掺杂在当代人类当中。 最近关于尼安德塔人 与丹尼索瓦人 的基因体排序也显示确实发生某些混杂。在非洲以外的现代人类在其基因体中,有2-4%的尼安德塔人的等位基因,美拉尼西亚人额外具有4-6%的丹尼索瓦人等位基因。这些新的结果并不与《源出非洲》模型相抵触,但显示发生了一些额外的混杂事件。可能源自于距今73,000年前的多峇超级火山灾难的一个种群瓶颈之后,经过一段复苏时间,一支很小的群体离开非洲,并遇到尼安德塔人,可能是在中东,他们的后代扩散到世界各地,有一小支遇到了丹尼索瓦人,可能是在东南亚,就在他们迁入美拉尼西亚之前。
参见
进化论
被禁止的历史
人类演化历程
早期人类对火的使用
巨人
水猿假说
免责声明:以上内容版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。感谢每一位辛勤著写的作者,感谢每一位的分享。

相关资料





- 有价值
- 一般般
- 没价值















推荐阅读




关于我们

APP下载


